Структурные элементы экосистемы
Для целей математического моделирования функции экосистемы необходимо максимально формализовать структурные и функциональные параметры экосистемы. Структура экосистемы складывается из структурных элементов фитоценоза и педоценоза. Она представлена трехмерным анизотропным пространством. По вертикальной оси можно выделить ярусы растительного покрова и горизонты почвенного профиля. По горизонтальной оси можно выделить контуры, выделы, парцеллы почвенного и растительного покрова, которые характеризуют горизонтальные размеры экосистемы.
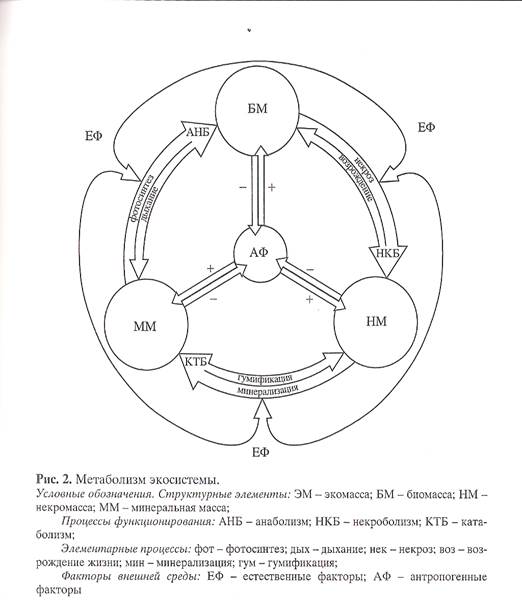
Объем экосистемы характеризуется общей массой ее вещества - экомассой имеющей анизотропную структуру. Она состоит из трех компонентов: биомассы (фитомасса, зоомасса, микробиомасса), некромассы (опад, подстилка, гумус), минеральной массы (газы, соли, коллоиды) (Рис. 2). Все компоненты экомассы также имеют анизотропную структуру с разным изменением параметров в вертикальном и горизонтальном направлениях.
Экосистема, как было сказано выше, представляет собой симбиотическое сообщество автотрофных и гетеротрофных организмов,
осуществляющее общую функцию метаболизма за счет взаимного обмена продуктами жизнедеятельности в определенном диапазоне гидротермических условий. Метаболизм экосистемы можно представить себе как биологический круговорот вещества между фитоценозом и педоценозом. С зооценозом немного сложнее.
С одной стороны, зооценоз можно отнести к педоценозу, поскольку он вместе с остальной гетеротрофной биотой трансформирует фитомассу и зоомассу в некромассу.
С другой стороны, зооценоз вполне можно представить как часть фитоценоза, выполняющего функцию вторичного анаболизма - синтеза зоомассы из фитомассы. В таком случае функцию анаболизма можно значительно расширить, поскольку ее участником становится вся биота экосистемы, без исключения, до тех пор, пока она живет и функционирует, т.е. синтезирует биомассу. Тогда анаболизм можно определить как взаимодействие биосинтеза и экскреции (все выделения биоты).
Экосистема как объект управления представляет собой экомассу, выполняющую функцию метаболизма. Эком асса состоитиз трех компонентов: биомасса, некромасса, минеральная масса, объем которых имеет характерную вертикальную и горизонтальную структуру.
Биомасса- совокупность фитомассы, зоомассы и микробиомассы. Фитомасса - первичная биологическая продукция, синтеЗИР9ванная фитоценозом и трансформированная затем зооценозом в зоомассу, а микробиотой в микробиомассу. Конечной продукцией анаболизма является биомасса экосистемы, а не только одна фитомасса. Хотя общая продукция биоценоза, включая выделения биоты, приурочена к надземным и подземным ярусам фитоценоза.
После завершении биологического цикла вся биота экосистемы без исключения, в соответствии с индивидуальной программой; онтогенеза и функции некроболизма с учетом факторов внешнего воздействия, отмирает и переходит в другое качественное состояние - в некромассу. Предварительно она переводит часть синтезированного вещества в зародыши будущих организмов.
Некромасса- отмершая биомасса в результате процесса катаболизма экосистемы (минерализация минус гумификация) трансформируется в минеральную массу, которая используется фитоценозом в процессе анаболизма, частично связываются в форме гумусовых соединений, накапливаясь в почве как стратегический запас элементов минерального питания. Не востребованные фитоценозом и не связанные гумусом минеральные элементы
частично вымываются из экосистемы водными потоками, а остальные образуют нерастворимые минеральные коллоиды (вторичные и первичные минералы) и осаждаются в почве.
В составе некромассы можно выделить три группы веществ:
- первичная некромасса: опад, отпад, валеж, ветошь, торф;
- вторичная некромасса: подстилка, гумус, сапропель,
- третичная некромасса: ППК, конкреции карбонатные и железомарганцевые, вторичные (глинистые) и первичные, , синтезированные в почве минералы.
Почва или педоценоз представляет собой совокупность гетеротрофной биоты, различных фракций некромассы и разнообразных продуктов ее минерализации и гумификации. Это запасной фонд экосистемы - ставший в процессе эволюции основным и I единственным источником элементов минерального питания для высших растений, Т.е. фитоценоза. Структура педоценоза представлена профилем с набором генетических горизонтов, а функция - процессом, который с помощью гетеротрофной биоты осуществляет минерализацию и гумификацию отмершей биомассы.
Минеральная масса - совокупность продуктов минерализации. некромассы в форме газов, солей и коллоидов. Газы и соли (ионы) поглощаются фитоценозом в процессе анаболизма, а коллоиды постепенно накапливаются в почве в форме вторичных и первичных минералов и создают минеральную основу почвенного профиля, на которой функционирует педоценоз.
Минеральная масса, в отличие от биомассы и некромассы, не имеет физического носителя. Эта масса в свободном состоянии бывает очень короткое время, когда перемещается из одного функционального блока в другой. Только отходы метаболизма в виде минеральных "шлаков", газовых эмиссий и солевого стока можно измерить и оценить количественно, поскольку они поддерживают стабильный состав атмосферы, гидросферы и литосферы. Тем не менее, количество минеральной массы в структурных элементах экосистемы можно измерить с помощью химического анализа проб почвы, биомассы, воздуха и лизиметрических вод.
1.3. Функция экосистемы. Функциональные блоки. Механизм функционирования
Для живых природных систем всех уровней организации характерен один универсальный механизм функционирования - обмен вещества и энергии или метаболизм. Классическая биология трактует метаболизм как способ проявления и поддержания жизни путем взаимодействия двух противоположно направленных
процессов: анаболизма и катаболизма. Анаболизм - это процесс ассимиляции простых (минеральных) веществ в сложные (органические). Катаболизм - противоположный процесс диссимиляции сложных (органических) веществ на простые (минеральные).
Метаболизма живых систем достаточно хорошо изучен на уровне клетки и организма. На более высоких уровнях организации живых систем (экосистема биом, биосфера) принято изучать циклы отдельных элементов (углерод, азот, сера и другие). В реальных экосистемах происходит регулярный обмен всего набора химических элементов, входящих в состав биоты. У каждого элемента своя траектория и характерное время круговорота. Для того, чтобы понять законы, управляющие круговоротом химических элементов и достоверно прогнозировать их поведение, необходимо
изучить механизм функционирования экосистем, его движущие силы и характер проявления в разных биоклиматических условиях.
Для осуществления функции метаболизма природная экосистема имеет не два (как клетка и организм), а три функциональных блока:
- блок анаболизма;
- блок некроболизма;
- блок катаболизма.
По сравнению с классическим определением метаболизма
клетки и организма как взаимодействия процессов анаболизма и катаболизма, на уровне экосистемы выделен третий функциональный блок - некроболизм,который осуществляет превращения живой биомассы в мертвую некромассу (Рис. 2).
Процесс некроболизма закодирован в генетической памяти каждого вида биоты. Он представляет собой вторую половину жизни каждого организма и начинается с момента включения генеративной фазы онтогенеза. К этому времени полностью сформированный организм способен добывать из внешней среды больше ресурсов, чем нужно для его собственного жизнеобеспечения. Этот избыток он направляет на зарождение новой жизни, на формирование потомства.
у растений генеративная фаза наступает при достижении определенной массы. После этого начинается массовое цветение, образование плодов и семян с закодированной программой развития будущего растения и минимальным количеством энергетического ресурса (эндосперма), необходимого для выведения к солнцу собственной антенны - проростка зеленого листа.
У животного, при достижении генеративного возраста, начинаются брачные игры, рождение, воспитание и обучение потомства. До наступления генеративной фазы организм работает на себя, обеспечивает собственное развитие до полного формирования морфологической структуры, заложенной в генетической памяти вида. С момента наступления генеративной фазы организм начинает функционировать ради будущего, постепенно передавая собственную жизнеспособность будущему поколению. Поэтому функция некроболизма - это не только заключительная фаза завершения жизни (некроз) отработавшего ресурс организма, но и начальная фаза возрождения новой жизни, а также сопровождение ее на первом беззащитном этапе развития.
Каждый блок представляет взаимодействие двух противоположно направленных процесса. Блок анаболизма представляет взаимодействие процессов фотосинтеза и дыхания, а в общем случае - биосинтеза и экскреций. Блок некроболизма – взаимодейсвие
процессов некроза и возрождение жизни. Блок катаболизма - взаимодействие процессов минерализации и гумификации некромассы.
По существу биологический круговорот или обмен вещества и энергии в экосистеме происходит между живой и мертвой биомассой. Однако фитоценоз в процессе анаболизма способен принимать от педоценоза только минеральные вещества в форме газов и растворов солей (ионов), высвобождаемые в процессе катаболизма. Поэтому в структуре экосистемы, кроме живой биомассы и мертвой биомассы (некромассы), мы сочли необходимым выделить минеральную массу как предмет обмена вещества между педоценозом (почвой) и фитоценозом (растительностью) .
Вся сложная совокупность вещества экосистемы составляет ее общую массу - экомассу, которая в процессе метаболизма экосистемы находится в постоянном движении, превращении, обновлении.
Процесс обновления экомассы каждой экосистемы идет с определенной скоростью, которая зависит от гидротермических условий экотопа и видового состава биоты, адаптированной к этим условиям. Время полного обновления экомассы называется характерным временем (ХВ) экосистемы. Оно измеряется временем обновления (существования) самой устойчивой фракции экомассы. В одних случаях самой устойчивой может оказаться фитомасса (лесные экосистемы), в других случаях - некромасса (степные, болотные экосистемы), в третьих - минеральная масса (солончаки, такыры). Характерное время отдельных фракций экомассы измеряется временем жизни самой устойчивой части этих фракций. Например, жизненный цикл популяции вида эдификатора определяет ХВ фитоценоза, цикл синтеза и распада гумуса определяет ХВ педоценоза (почвы), геохимический цикл биогенных элементов определяет ХВ экосистемы.
Характерные времена (ХВ) самых разных экосистем биосферы измеряются довольно близкими величинами в пределах300-500 лет. За это время происходит периодическое обновление фитомассы популяции эдификаторов лесных экосистем, некромассы степных и болотных экосистем, солевого состава солончаков и других засоленных почв. Если же ХВ экосистем измерять по времени обновления некромассы, то для подзолистых почв оно составит 60-70 лет, для серых лесных -120-140 лет, для черноземов – 400-500 лет, для красноземов - 5-10 лет. Поэтому в любом гидротермическом диапазоне экосистемы функционируют в сходном ритме и в стационарном режиме, который согласуется с солнечно-земными ритмами.
Великое разнообразие живых существ с разными временными циклами онтогенеза (от 10 дней до 1000 лет), такое же разнообразие фракций некромассы с разными сроками жизни (от нескольких минут лизиса клеток до 1000 лет существования в почвах гуминов) создали гармонию взаимодействия живого и мертвого вещества экосистемы. Благодаря этому: взаимодействию, экосистема функционирует практически автономно без привлечения ресурсов геологического круговорота: атмосферы, гидросферы, литосферы.
Для автономного функционирования экосистем нужен только приток солнечной энергии и периодическая компенсация перехода части вещества из биологического в геологический круговорот. Солнце пока надежно освещает Землю, а потери экосистем в геологический круговорот вполне компенсируются за счет космических выпадений, согласно данным С.П. Голенецкого (Гoленецкий, Жигаловская, Голенецкая. 1981; Голенецкий, Малахов, Степанюк, 1981; Голенецкий, Степанюк, 1983). Поэтому не стоит однозначно говорить о том, что биосфера существует за счет ресурсов атмосферы, гидросферы, литосферы. Вполне возможно предположить обратное: метаболизм биосферы поддерживает их постоянный состав и динамическое равновесие.
Концепция экосистемы как информационно-управляющей системы была разработана при непосредственном участии В.А. Ковды (Ковда, Бугровский, Керженцев, Зеленская, 1990) и.
апробирована в ходе международного эксперимента "Убсу-Нур" под руководством В.В. Бугровского. Этот, по существу, подготовительный этап работы занял очень много времени(1980-1995 гг.). Очень трудно было отрешиться от привычных представлений естественных наук и усвоить методологию и терминологию наук технического профиля. Это был принципиальный переход от изучения структуры экосистем к изучению их функций, равный по значимости переходу от анатомии к физиологии организма.
Главная сложность перехода заключалась в том, что традиционно все компоненты экосистемы и все факторы внешней среды изучались раздельно, как бы независимо друг от друга, самостоятельными научными дисциплинами. Параметры каждого компонента считались уникальными и измерялись разными методами в разных единицах, оценивались по разным не согласованным критериям. Это оказалось серьезным препятствием при интегрировании отдельных компонентов в единую и целостную экосистему. Пришлось искать такую позицию, такую точку видения объекта, из которой все компоненты представляют функциональное единство, а их уникальность проявляется только в той
роли, которую они выполняют в едином процессе метаболизма экосистемы, в ее главной функции. Именнр функция метаболизма экосистемы оказалась таким объединяющим началом. .
Схема, представленная на рис. 2 отражает наше понимание структуры экосистемы и механизма ее функционирования; связи экосистемы с естественными и антропогенными факторами внешней среды. Многие процессы, термины и механизмы, указанные в схеме, давно известны в общей и молекулярной биологии, широко используются на уровне клетки и организма. Наша задача состояла в том, чтобы перенести их на уровень экосистемы. При этом мы учитывали принципиальную разницу между уровнем организма и экосистемы, которая заключается в том, что при изменении внешних условий организм стремится сохранить структуру путем изменения режима функционирования, 'аэкосистема наоборот, манипулирует структурой ради сохранения функции.
Метаболизм экосистемы представляет собой необратимый циклический I{pyroBopoT вещества между фитоценозом и педоценозом.Метаболизм экосистемы включает в себя три последовательных процесса преобразования, ее массы - экомассы:
- анаболизм (фотосинтез-дыхание) - превращает минеральную массу в фитомассу с помощью солнечной энергии, часть фитомассы превращается в зоомассу и микробиомассу;
- некроболизм (некроз-возрождение) - превращает биомассу в некромассу при этом часть вещества направляется в генеративные органы на возрождение потомства;
- катаболизм (минерализация-гумификация) - превращает некромассу в минеральную массу, при этом часть минеральной массы, взаимодействует с продуктами полураспада, образуя почвенный гумус, выполняющий в экосистеме одновременно три функции: накопителя; хранителя и дозатора элементов минерального питания фитоценоза.
В процессе анаболизма или ассимиляции простых минеральных веществ в сложные органические участвует вся биота экосистемы, но инициатором этого главного процесса в метаболизме экосистемы, становится фитоценоз - сообщество автотрофных организмов. Автотрофные организмы (продуценты) обладают способностью превращать минеральные вещества в органические с помощью солнечной энергии. Они I создают первичную биологическую продукцию. Гетеротрофные организмы (консументы и редуценты) преобразуют первичную биологическую продукцию (фитомассу) во вторичную (зоомассу и микробиомассу), используя для этого энергию химических связей, накопленную
продуцентами в фитомассе. Совокупный продукт анаболизма - суммарная биомасса экосистемы, состоит из фитомассы, зоомассы и микробиомассы. В процессе анаболизма происходит взаимодействие двух противоположных процессов: биосинтеза (biosyпthesis) и экскреции (excretioп). Это означает, что часть синтезированной биомассы расходуется на жизнеобеспечение самих организмов (продуцентов, консументов и редуцентов).
Некроболизм - процесс превращения биомассы в некромассу вследствие естественного завершения жизненного цикла любого организма. Этот процесс запрограммирован в генетическом аппарате всех живых существ без исключения. Он начинается сразу после достижения организмом генеративной фазы развития, когда значительная часть продуктов анаболизма направляется на создание и поддержание новой жизни, а не только на собственное жизнеобеспечение, как в период вегетативного развития. Полностью морфологически сформированный организм способен добывать гораздо больше ресурсов, чем необходимо для его собственного жизнеобеспечения. Вот почему задолго до отмирания растения его жизненно важные ассимиляты перемещаются в органы и ткани, обладающие потенциалом функционирования (запасающие органы, цветы, плоды, семена). У животных в этой фазе происходит спаривание, вынашивание плода, воспитание потомства, передача ему навыков охоты и защиты, строительства или поиска жилища, хранения запасов пищи и другое.
В экосистеме некроболизм выполняет своеобразную буферную функцию, которая позволяет процессам анаболизма и катаболизма гармонично взаимодействовать между собой при флуктуации факторов внешней среды. Некроболизм также как и анаболизм нельзя отождествлять только с одним процессом отмирания, поскольку он складывается из двух противоположных процессов: отмирания (пecroze) и возврата полезных для потомства ресурсов в запасающие и генеративные органы, то есть возрождения жизни (revival). В процессе отмирания отработавшей жизненный ресурс биомассы, происходит вторичный синтез зародышей новой будущей биомассы. Именно здесь заложен ключевой момент материального обеспечения продолжения жизни в механизме функционирования экосистемы.
Функцию катаболизма или диссимиляции сложных органических веществ в простые минеральные выполняет педоценоз - сообщество гетеротрофных организмов. Процесс превращения некромассы в минеральную массу складывается из двух противоположно направленных процессов: минерализации и гумификации - вторичного синтеза органического вещества (почвенного гумуса). В метаболизме экосистемы минерализация некромассы
обеспечивает снабжение фитоценоза элементами минерального питания (ЭМП) для осуществления им функции анаболизма. Гумификация выполняет сразу три роли: накопителя, хранителя и дозатора ЭМП для растений. Эти три роли в метаболизме экосистемы играет почва, а точнее педоценоз - сообщество гетеротрофных организмов вместе с исходными, конечными и промежуточными продуктами деструкции некромассы.
Процесс функционирования экосистемы в самом общем виде представляет собой последовательное превращение биомассы в некромассу, некромассы в минеральную массу, минеральной массы в биомассу с помощью процессов анаболизма, некроболизма и катаболизма. Разнообразные организмы с различными характерными временами онтогенеза, разной ритмикой и продолжительностью жизни (от нескольких дней до нескольких столетий) создают сложную, многослойную, цикличную и одно направленную систему непрерывного процесса метаболизма экосистемы ее главной функции.
Величина (масса) каждого структурно-функционального блока экосистемы зависит от соотношения скоростей противоположных (входных и выходных) процессов. Например, величина биомассы представляет собой разницу между результативностью процессов анаболизма и некроболизма; некромасса - это результат преобладания результативности процесса некроболизма над результативностью катаболизма. Величина минеральной массы находится в прямой зависимости от соотношения результатов катаболизма и анаболизма. Интенсивность всех трех процессов составляет режим функционирования экосистемы первого уровня, который формирует и регулирует общую структуру экосистемы, поддерживает ее морфологическое строение и диагностические признаки, позволяющие идентифицировать данную экосистему, отличить ее от других подобных.
Второй уровень функционирования составляют более частные процессы. Эффективность анаболизма, например, определяется соотношением интенсивности биосинтеза (фотосинтеза) и выделения (дыхания); эффективность некроболизма зависит от соотношения скорости отмирания и возврата ассимилятов из отмирающих органов и организмов в функционирующие, в том числе и в генеративные; эффективность катаболизма определяется соотношением скорости минерализации некромассы и вторичного синтеза почвенного гумуса или интенсивности процесса гумификации.
Регуляторами всех частных процессов функционирования и общего мутаболизма экосистемы являются факторы внешней среды, которые принято разделять на естественные, антропогенные
и смешанные. Важнейшие естественные факторы среды: свет, тепло и влага. Главными антропогенными факторами можно считать: изъятие, привнос и трансформацию массы вещества каждого структурного блока экосистемы. К смешанным факторам следует относить искусственное изменение света, тепла и увлажнения, а также естественное изъятие, привнос и трансформацию массы вещества в результате жизнедеятельности биоты, а также вследствие стихийных катастроф и других нестандартных аномальных явлений.
Воздействия естественных, антропогенных и смешанных факторов имеют принципиально разные последствия для структуры и функции экосистем. Естественные факторы оказывают непосредственное влияние на процессы функционирования, стимулируя или сдерживая их интенсивность, а через них опосредованно эти факторы влияют на величину массы экосистемы и ее структурных блоков. Антропогенные факторы оказывают прямое воздействие на величину массы экосистемы и ее структурных блоков, не затрагивая интенсивность процессов или, оказывая на них опосредованное влияние, через изменение массы (вырубка леса, сбор урожая, внесение органических и минеральных удобрений, выпас скота и т.п.).
Иногда естественные процессы оказывают прямое воздействие на массу экосистемы: пожары, инвазии энтомовредителей, землетрясения, наводнения, обвалы, оползни и другие стихийные бедствия. Некоторые антропогенные факторы оказывают прямое воздействие на процессы функционирования экосистем: распашка земель, орошение, осушение, мульчирование и другие факторы, стимулирующие или сдерживающие активность метаболизма экосистемы. Искусственно измененные скорости процессов могут, в свою очередь, изменить массу вещества того или иного структурного блока экосистемы. Именно таким способом происходит стимулирование урожаиности сельскохозяиственных культур, увеличение или уменьшение запаса гумуса в почве и другое.
Из сказанного выше следует, что колебания естественных факторов, даже экстремальные, не способны резко изменить структуру экосистемы, поскольку для этого надо изменить соотношение скоростей сразу нескольких противоположно направленных процессов. Изменившиеся условия сначала изменяют скорость функционирования, а потом измененные скорости функционирования изменяют прежнюю структуру на новую, более адаптированную к новым условиям. Эта новая структура позволяет экосистеме успешно функционировать в изменившихся условиях внешней среды с прежней эффективностью.
Для восстановления нарушенного равновесия после возврата прежних условий бывает достаточно короткопериодных сукцессий, иногда даже без существенной смены видового состава. При длительном отклонении факторов среды от оптимального диапазона происходит метаморфоз экосистемы, то есть переход ее изодного таксона классификации в другой. Долговременные флуктуации условий среды в устоявшихся границах оптимального диапазона сопровождаются уплотнением экологических ниш и повышением КПД экотопа, что приводит к образованию новых видов и формированию новых типов экосистем, то есть их эволюционным изменениям.
Воздействие антропогенных факторов способно резко изменить структуру экосистемы, поскольку они оказывают прямое воздействие на ее массу (привнос, вынос, трансформация). Антропогенные факторы по силе воздействия сопоставимы со стихийными природными бедствиями, компенсировать последствия которых может только длительная экзогенная сукцессия со сложной многократной и длительной сменой видового состава(зарастание вырубки, гари и залежи).
Смешанные факторы могут оказывать прямое воздействие как на функции, так и на структуру экосистемы. Культура закрытого грунта, где искусственно регулируется весь набор естественных факторов среды: свет, тепло, влага, - обеспечивает получение максимальной биомассы или наиболее полезной для человека ее части. Примером противоположного аспекта может служить уничтожение фитомассы в результате инвазии саранчи или сибирского шелкопряда, которые сопровождаются сменой микроклимата и соответствующими изменениями функций экосистемы с последующей длительной экзогенной сукцессией со сложной последовательной сменой фитоценозов.
Метод технико-биологических аналогийпозволяет решать не только философские, но и конкретные вопросы функциональной экологии, например, управления функциями природных экосистем. Именно этот метод дает практическую возможность изучать природные системы как объекты управления.
Управление функциями природных систем необходимо для сохранения, восстановления и поддержания экологического равновесия в условиях нарастающего антропогенного воздействия. Процесс управления представляет собой серию заранее рассчитанных воздействий, направленных на повышение продуктивности природных систем и их устойчивости к негативным воздействиям естественных и антропогенных факторов.
В расчеты ожидаемых результатов от процедуры управления должны закладываться не только максимальный эффект для человека,
но и минимальный ущерб для природы. Добиваясь конкретных полезных результатов, нельзя допускать возможности вредных последствий для экосистемы даже в отдаленной перспективе. Опыт показал, что затраты на исправление допущенных ошибок, как правило, многократно превышают первоначальную выгоду.
Иными словами, направленно воздействовать на механизм функционирования экосистем желательно в пределах естественного диапазона его изменчивости. Для решения этой задачи нужно изучить устройство и принцип действия механизма функционирования экосистем, возможные реакции этого механизма на воздействия внешних факторов, пределы допустимых воздействий и критических изменений параметров экосистем, после которых начинаются нарушения их структуры и функций. Нужна четко сформулированная и строго аргументированная теория управления механизмом функционирования экосистем, которая позволила бы прогнозировать их поведение при тех или иных изменениях факторов внешнего воздействия.
Сложный и длительный процесс разработки теории управления функциями природных систем можно значительно облегчить и ускорить за счет использования опыта, накопленного техническими науками в области управления сложными техническими системами. Основная трудность такого подхода, несмотря на его привлекательность, заключается в том, чтобы доказать сходство или хотя бы подобие между природными и техническими системами, которые принято считать антиподами.
Мы упоминали выше принцип информационного единства природных и технических систем, который позволяет обнаружить черты сходства в процессе функционирования тех и других систем. Суть его заключается в том, что природные и технические системы имеют не только вещественную, но и информационную форму существования. В технических системах эти формы представляют - машина и ее чертежи или техническая документация, в природных системах - фенотип и генотип или организм и семя, которое превратится в новый организм. Для того, чтобы информационные формы трансформировались в вещественные необходимо приложить к ним вещество и энергию.
Чертежи машин не воплощенные в металле, невсхожие семена растений, погибшие зародыши животных - это примеры некоторой автономии информационных форм существования природных и технических систем по отношению к вещественным. Несмотря на то, что вещественные формы существования природных и технических систем сильно различаются между собой, их информационные формы имеют много общего и вполне сопоставимы.
Обе эти формы можно отнести к одному классу информационноуправляющих систем. Разница между ними заключается в том, что механизм действия и алгоритм поведения технических систем создан и заложен в систему волей и действиями человека, а механизм управления природными системами отработан в течение длительного процесса эволюции Жизни и содержится в генетической программе развития каждого организма. Программа управления жизнью организма заложена в семени и ее реализация зависит от соответствия идеальных и реальных условий среды. Это означает, что информация о динамике факторов среды должна соответствовать программе онтогенеза организма, заложенной в семени. Отклонение условий в лучшую или худшую сторону от оптимума на каждом этапе развития может ускорить или замедлить ход физиологических процессов и изменить результативность онтогенеза.
Условия, при которых происходит переход информационной формы в вещественную, могут значительно отличаться от идеальных как у технических, так и у природных систем. Отклонение технических условий от заданных в проектной документации, отражается на качестве технических систем, их надежности, работоспособности, долговечности. Отклонение параметров среды и динамики их изменчивости от оптимального диапазона отражается на структуре и функциях природных систем, на итогах онтогенеза и каждой фенофазы. Следствием этого может стать сохранение, увеличение или уменьшение конкурентоспособности организма в ответ на изменения факторов среды. В результате организм может либо сохранить свои позиции в экосистеме, либо усилить их и расширить ареал, либо ослабить и выпасть из экосистемы.
Механизм функционирования природных системвсех уровней организации (клетка, организм, экосистема, биом, биосфера) - метаболизм принципиально не различается. Круговорот вещества. И энергии совершается примерно одинаково на всех уровнях организации живых систем и представляет многократную циклическую смену процессов синтеза и распада биомассы. Различия механизма' функционирования разных уровней организации природных систем касаются только количественной стороны, а принцип действия идентичен: анаболизм, некроболизм, катаболизм.
Все природные системы развиваются на фоне флуктуации факторов среды (суточных, сезонных, годовых, многолетних) в режиме непрерывной адаптации к этим постоянным изменениям. Каждая природная система имеет свой диапазон оптимальных флуктуаций, заложенный в генетическом коде. Отклонение реального
хода смены условий от идеального, генетически обусловленного, сказывается на конечном итоге развития системы. Аномалии проявляются в изменении габитуса, массы, продукции системы. Выход условий среды за пределы оптимального диапазона может блокировать генеративную фазу онтогенеза и привести к гибели вида. Отдельные мутанты, выжившие в экстремальных условиях, могут стать родоначальниками нового вида, адаптированного к данным условиям.
Маятниковый характер флуктуации факторов среды приводит к тому, что каждая природная система вынуждена попеременно реагировать то на улучшение, то на ухудшение условий среды, поскольку оптимальные условия каждой системы находятся между кардинальными точками min и mах. Точка min означает состояние анабиоза системы, а точка mаx - ее предельные возможности.
Реакцию механизма функционирования экосистем на изменения факторов среды можно сравнить с работой двигателя внутреннего сгорания на разных оборотах. Наибольшая эффективность и долговечность работы двигателя достигается на средних оборотах в оптимальном режиме. При низких оборотах двигатель не достигает реальной мощности, а при высоких оборотах он работает на износ с перерасходом ресурсов.
У природных систем также существует некоторая разница реакции на отклонения факторов среды от оптимума в сторону min и mах. При улучшении условий (mах) адаптивная реакция системы не встречает внутреннего сопротивления, мобилизует все возможности организма, вследствие чего эффект получается позитивный и адекватный улучшению условий. При ухудшении условий среды (min) живая система включает буферные механизмы, сдерживающие ее реакцию на неблагоприятную смену условий среды, что снижает общую эффективность функционирования в меньшей степени, чем того требуют условия. Поэтому после прохождения полного синусоидального цикла флуктуации условий (суточного, годового, многолетнего) практически всегда получается положительный эффект, который выражается в некотором увеличении биомассы, урожая плодов и семян.
Следовательно, при любых сочетаниях условий среды любая природная система развивается по наиболее выгодной для нее траектории. Видимо на этой принципиальной основе происходит процесс экспансии Жизни, непрерывного увеличения биомассы Земли и на ней же базируется предсказание В.И. Вернадского о растекании Жизни по Земле, ближнему и дальнему Космосу.
Функционирование информационно-управляющих систем должно быть подчинено определенной цели, для достижения которой
они имеют набор функциональных блоков, уровней управления и соответствующих уровней информации, корректирующих процесс управления. Целесообразность технических систем определяется человеком, поскольку всякая машина предназначена для какой-то цели; средством достижения которой она и является. У природных систем цель заложена в генотипе. Эту цель можно сформулировать как "воспроизводство себе подобных". Для достижения этой цели служит "конструкция" или структура природной экосистемы, которая состоит из функциональных блоков. Функционирование каждого блока определяется уровнями управления, которые контролируются соответствующими уровнями информации о динамике факторов среды и состоянии соседних блоков системы.
Ради воспроизводства себе подобных (репродукции информационных форм) живые системы адаптируются в пределах генотипа к меняющимся условиям среды. В зависимости от характера и степени отклонения реальной среды от идеальной, обусловленной генетическим кодом, живые системы могут изменить ритмику функционирования, массу и габитус. Однако су<
Дата добавления: 2020-10-01; просмотров: 938;
Поиск по сайту
Узнать еще
- IV.3. Элементы стратегии выживания человечества
- VI.II. Элементы складки
- А - решетчатая конструкция из бетонных элементов; б - пространственная георешетка; в - укрепление откоса георешеткой; 1, 2 - бетонные элементы; 3 - анкеры; 4 - тяжи анкеров
- Активные и пассивные элементы электрических цепей. Закон Ома
- Анализ возможных последствий неадекватного поведения элемента на смежные элементы
- Антропогенное воздействие на экосистемы
- Архитектурные элементы
- Б. Элементы контрольных приспособлений

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
