Типы корней и корневых систем
По происхождению корни делятся на главные, боковые и придаточ- ные. Главный корень семенных растений развивается из корешка зародыша семени. Стебель является продолжением корня, и вместе они составляют ось 1-го порядка. Место сочленения оси и семядольных листьев называют семядольным узлом. Участок, расположенный на границе главного корня и стебля, именуется корневой шейкой. Участок стебля от корневой шейки до первых зародышевых листьев (семядолей) называют подсемядольным коленом, или гипокотилем, а от семядолей до первых настоящих листьев - эпикотилем, или надсемядольным коленом. У двудольных и голосеменных растений от главного корня за счет меристематической активности перицикла отходят боковые корни 1-го порядка, которые дают начало боковым корням 2-го и 3-го порядка. Корневая система, образованная системой главного корня, называется стержневой, а с развитой системой боковых корней - ветвистой; таким образом, ветвистая корневая система является разновидностью стержневой. Чем больше боковых корней отходит от главного, тем больше площадь питания растения.
У большинства двудольных растений главный корень сохраняется всю жизнь, у однодольных растений главный корень не развивается, так как зародышевый корешок быстро отмирает, а от базальной части побега берут начало придаточные корни. Придаточные корни могут образовываться от листьев, стеблей, старых корней и даже от цветков и иметь ответвления 1-го, 2-го порядка и т.д. Корневая система, образованная придаточными корнями, называется мочковатой (рис. 3.2).
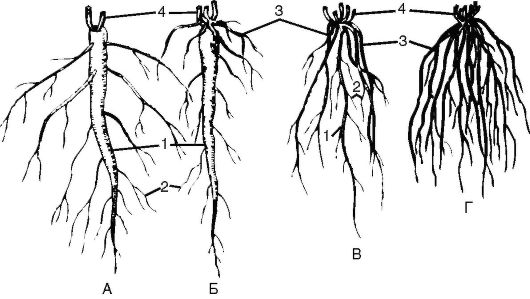
Рис. 3.2.Типы корневых систем: по форме: А, Б - стержневая; В, Г - мочковатая; по происхождению: А - система главного корня; Б, В - смешанная корневая система; Г - придаточная корневая система; 1 - главный корень; 2 - боковые корни; 3 - придаточные корни; 4 - основания побегов
У многих двудольных корневищных растений главный корень часто отмирает и преобладает система придаточных корней, отходящих от корневища (лютик ползучий, сныть обыкновенная).
По отношению к субстрату корни бывают следующих типов: земляные - развиваются в почве; водные - находятся в воде (у плавающих водных растений); воздушные, развивающиеся в воздушной среде (у растений, имеющих корни на стволах и листьях).
Зоны корня
В молодом корне различают 4 зоны: деления, растяжения, всасывания, проведения (рис. 3.3).
К зоне деления относят верхушку конуса нарастания (протяженность менее 1 мм), где происходит активное митотическое деление.
Верхушечная меристема откладывает наружу клетки корневого чехлика, а внутрь - ткани остальной части корня. Эта зона состоит из тонкостенных паренхимных клеток первичной меристемы, которые прикрыты корневым чехликом, выполняющим защитную функцию при продвижении корня между частичками почвы. От соприкосновения с почвой клетки чехлика постоянно разрушаются, образуя слизь, которая предохраняет зону деления при трении о по- чву и продвижении корня вглубь. У большинства растений корневой чехлик восстанавливается за счет первичной меристемы, а у злаков - за счет особой меристемы калиптрогена.
По теории гистогенов (Ганштейн, 1868), у большинства покрытосеменных растений апикальные меристемы состоят из 3 гистогенных слоев, различающихся направлением деления клеток и имеющих по 1-4 инициальные клетки. Самый наружный слой - дерматоген - формирует протодерму, из которой образуются клетки корневого чехлика и ризодерма - первичная покровно-всасывающая ткань в зоне всасывания. Средний слой - периблема - дает начало всем тканям первичной коры. Третий слой инициалей формирует плером, из которого развиваются ткани центрального осевого цилиндра.
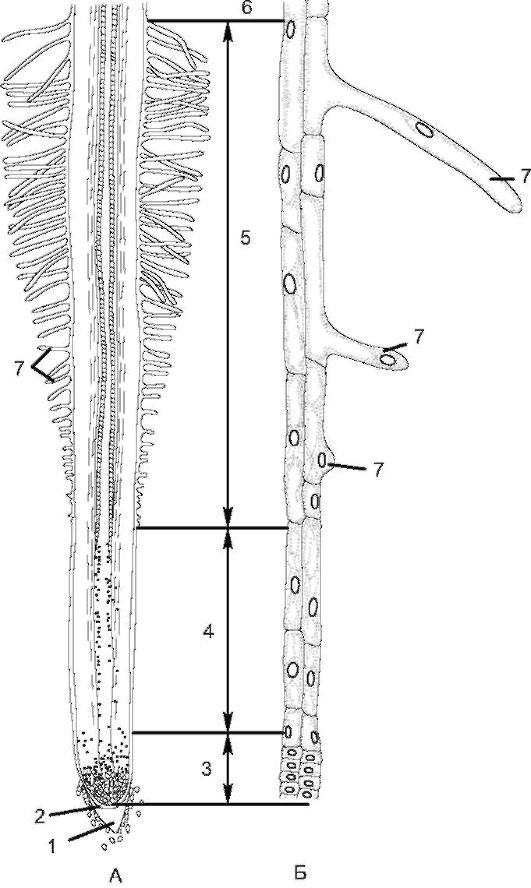
Рис. 3.3.Зоны корня (у проростка пшеницы): А - схема строения корня; Б - периферические клетки отдельных зон при большом увеличении: 1 - корневой чехлик; 2 - калиптроген; 3 - зона деления; 4 - зона растяжения; 5 - зона всасывания; 6 - зона проведения; 7 - корневые волоски клеток.
В зоне растяжения клетки меристемы увеличиваются в размерах (вследствие оводнения), вытягиваются в длину, и деление клеток постепенно прекращается. Вследствие вытягивания клеток в продольном направлении происходят рост корня в длину и его продвижение в почве. Зону деления и зону растяжения с учетом сохранения в них меристематической активности можно объединить в одну - зону роста. Ее протяженность составляет несколько миллиметров. В зоне всасывания происходит становление первичной структуры корня.
Протяженность зоны всасывания - от нескольких миллиметров до нескольких сантиметров; она характеризуется наличием корневых волосков, являющихся выростами клеток ризодермы. При их формировании ядро перемещается в переднюю часть корневого волоска. Последние увеличивают всасывающую поверхность корня и обеспечивают активное всасывание воды и растворов солей, но они недолговечны (живут 10-20 дней). Новые корневые волоски образуются под зоной всасывания, а отмирают - над этой зоной. По мере роста растения зона всасывания постепенно перемещается, и растение имеет возможность поглощать минеральные вещества из разных слоев почвы.
Постепенно зона всасывания переходит в зону проведения (укрепления). Она тянется вплоть до корневой шейки и составляет по протяженности большую часть корня. В этой зоне идет интенсивное ветвление главного корня и появляются боковые корни. У двудольных растений в зоне проведения формируется вторичная структура корня.
Анатомия корня
Первичное строение корня (рис. 3, см. цв. вкл.). Строение корня в зоне всасывания называется первичным, потому что здесь происходит дифференциация тканей из первичной меристемы конуса нарастания. Первичное строение корня в зоне всасывания можно наблюдать у двудольных и однодольных растений, но у однодольных оно сохраняется на протяжении всей жизни растения. На поперечном срезе корня первичного строения выделяют 3 основные части: покровно-всасывающую ткань, первичную кору и центральный осевой цилиндр (рис. 3.4).
Покровно-всасывающая ткань - ризодерма (эпиблема) выполняет как покровную функцию, так и функцию интенсивного всасывания воды и минеральных веществ из почвы. Клетки ризодермы живые, с тонкой целлюлозной стенкой. Из некоторых клеток ризодермы формируются корневые волоски; каждый из них представляет собой длинный вырост одной из клеток ризодермы, ядро же клетки обычно находится в кончике выроста. Корневой волосок содержит тонкий пристенный слой цитоплазмы, более плотный на верхушке волоска, а в центре - крупную вакуоль. Корневые волоски недолговечны и в зоне укрепления отмирают. Физиологически зона всасывания представляет собой очень важную часть корня. Клетки ризодермы поглощают водные растворы всей поверхностью наружных стенок. Развитие корневых волосков во много раз увеличивает поверхность поглощения. Протяженность зоны всасывания - от 1 до 1,5 см.
Со временем эпиблема может слущиваться, и тогда покровную функцию выполняет экзодерма, а после ее разрушения - слой кле- ток мезодермы и иногда мезодермы и перицикла, стенки которых опробковевают и одревесневают. Поэтому диаметр старых корней однодольных растений меньше, чем молодых.
Первичная кора корня развита более мощно, чем центральный осевой цилиндр. Она состоит из 3 слоев: экзодермы, мезодермы (рис. 4, см. цв. вкл.) (паренхима первичной коры) и эндодермы. Клетки экзодермы многоугольные по форме, плотно сомкнуты и расположены в несколько рядов. Клеточные стенки пропитаны суберином,
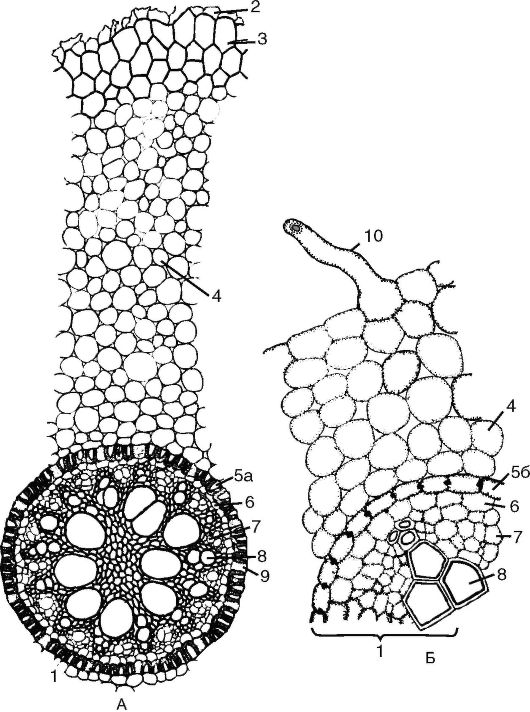
Рис. 3.4.Поперечный срез корня первичного строения: А - первичное строение корня однодольного;
Б - первичное строение корня двудольного: 1 - центральный (осевой) цилиндр; 2 - остатки эпиблемы; 3 - экзодерма; 4 - мезодерма; 5а - эндодерма с подковообразными утолщениями; 5б - эндодерма с поясками Каспари; 6 - перицикл; 7 - первичная флоэма; 8 - сосуды первичной ксилемы; 9 - пропускные клетки эндодермы; 10 - корневой волосок
т.е. опробковевают. Опробковение обеспечивает непроницаемость клеток для воды и газов. В экзодерме, обычно под корневыми волосками, сохраняются клетки с тонкими целлюлозными стенками - пропускные клетки, через которые проходят вода и минеральные вещества, по- глощенные ризодермой. Обычно они располагаются напротив лучей ксилемы радиального пучка.
Под экзодермой находятся живые паренхимные клетки мезодер- мы. Это наиболее широкая часть первичной коры. Клетки мезодермы выполняют запасающую функцию, а также функцию проведения воды и растворенных в ней солей от корневых волосков в центральный осевой цилиндр.
Внутренний однорядный слой первичной коры представлен эндодермой. Клетки эндодермы плотно сомкнуты и почти квадратные в поперечном сечении. В зависимости от степени утолщения клеточной стенки различают 2 типа эндодермы - с поясками Каспари (на поперечном срезе они выглядят как пятна Каспари) и с подковообразными утолщениями стенок.
Эндодерма с поясками Каспари - это начальный этап формирования эндодермы, при котором утолщению подвергаются лишь ее радиальные стенки за счет отложения веществ, сходных по химическому составу с суберином и лигнином. У многих двудольных и голосеменных растений процесс дифференциации эндодермы поясками Каспари заканчивается. У эндодермы с подковообразными утолщениями образуется толстая вторичная клеточная стенка, пропитанная суберином, в дальнейшем она одревесневает. Неутолщенной остается только наружная клеточная стенка (рис. 3.5). Эндодерма с подковообразными утолщениями развивается чаще у однодольных растений (рис. 5, см. цв. вкл.).
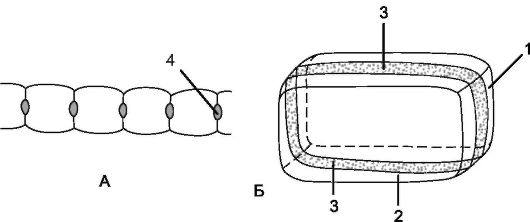
Рис. 3.5.Схема строения клетки эндодермы: А - общий вид; Б - поперечный разрез клеток: 1 - поперечная стенка клетки; 2 - продольная радиальная стенка; 3 - поясок Каспари; 4 - пятна Каспари
Считается, что эндодерма выполняет функцию гидравлического барьера, способствуя продвижению минеральных веществ и воды из первичной коры в центральный осевой цилиндр и препятствуя их выходу обратно.
Центральный осевой цилиндр начинается с клеток перицикла, который обычно в молодых корнях состоит из живых тонкостенных паренхимных клеток, расположенных в один ряд (но может быть и многослойным - например, у грецкого ореха). Клетки перицикла дольше других тканей корня сохраняют свойства меристемы и спо- собность к новообразованиям. Из перицикла образуются боковые корни, поэтому его называют корнеродным слоем. Проводящая система корня представлена одним радиальным сосудисто-волокнистым пучком, в котором группы элементов первичной ксилемы чередуются с участками первичной флоэмы. У однодольных растений количество лучей первичной ксилемы - 6 и более, у двудольных - от 1 до 5. Корни в отличие от стеблей не имеют сердцевины, так как в центре корня располагаются лучи первичной ксилемы.
Таблица 3.1.Формирование тканей корня первичного и вторичного строения
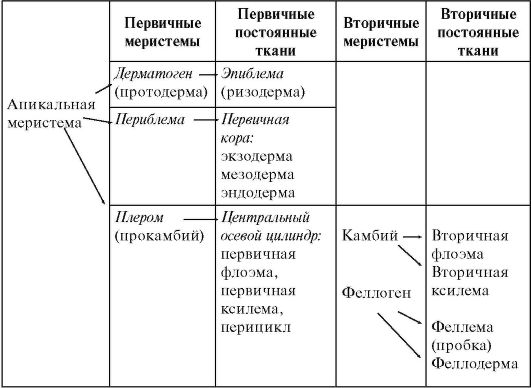
У однодольных и споровых архегониальных растений строение корня не претерпевает значительных изменений в течение всей жизни растения. У голосеменных и двудольных растений на границе зон всасывания и проведения происходит переход от первичного строения корня к вторичному (табл. 3.1).
Вторичное строение корня. В корнях голосеменных и двудольных растений камбий возникает из прокамбия (камбиальных дуг) за счет тангентального деления тонкостенных клеток, расположенных с внутренней стороны от флоэмных тяжей. На поперечном срезе клетки камбия представлены вогнутыми внутрь дугами (рис. 6, см. цв. вкл.). Клетки камбия образуют к центру вторичную ксилему (древесину), а к периферии - вторичную флоэму (луб). Вторичной ксилемы всегда бывает больше, чем вторичной флоэмы, и она оттесняет камбий наружу.
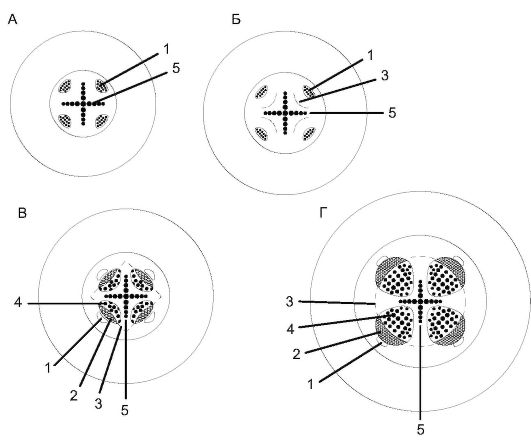
Рис. 3.6.Схема развития вторичного строения в корне: А - первичное строение; Б - заложение камбия; В - начало образования вторичных коллатеральных пучков; Г - вторичное строение корня: 1 - первичная флоэма; 2 - вторичная флоэма; 3 - камбий; 4 - вторичная ксилема; 5 - первичная ксилема
При этом дуги камбия сначала выпрямляются, а затем принимают выпуклую форму.
Когда дуги камбия достигают перицикла, его клетки тоже начинают делиться и образуют камбий межпучковый, а тот, в свою очередь, - сердцевинные лучи, представленные паренхимными клетка- ми, отходящими от лучей первичной ксилемы. Сердцевинные лучи, образованные межпучковым камбием, - это изначально «первичные лучи».
Таким образом, в результате деятельности камбия в корне между лучами первичной ксилемы формируются открытые коллатеральные сосудисто-волокнистые пучки, число которых равно числу лучей первичной ксилемы. Первичная флоэма при этом оттесняется вторичными тканями к периферии и сплющивается (рис. 3.6 и 3.7).
В перицикле, кроме межпучкового камбия, может закладываться феллоген, дающий начало перидерме - вторичной покровной ткани. При тангентальном делении клеток феллогена наружу отделяются клетки пробки, а внутрь - клетки феллодермы. Непроницаемость клеток пробки, пропитанных суберином, является причиной изоляции первичной коры от центрального осевого цилиндра. Первичная кора при этом постепенно отмирает и сбрасывается. Все ткани, располагающиеся от периферии до камбия, входят в понятие «вторич- ная кора» (рис. 7, см. цв. вкл.). В самом центре осевого цилиндра сохраняются лучи первичной ксилемы (от 1 до 5) (рис. 8, см. цв. вкл.), между которыми располагаются открытые коллатеральные пучки в количестве, соответствующем лучам первичной ксилемы (рис. 3.8).
Дата добавления: 2016-05-30; просмотров: 8211;
Поиск по сайту
Узнать еще
- Arthropoda. Клещи. Систематика. Морфология. Медицинское значение.
- Arthropoda. Паукообразные. Систематика. Географическое распространение. Морфология. Скорпионы. Пауки. Медицинское значение.
- Arthropoda..Систематика.Насекомые.Морфология.Классификация.Медицинское значение.
- Arthropoda.Систематика.Блохи.Виды блох.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское и эпидемиологическое значение.Меры борьбы.
- Arthropoda.Систематика.Мошки,мокрецы,слепни,оводы.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское значение,меры борьбы.
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- Cистеми числення і способи переведення чисел із однієї системи числення в іншу

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
