Вторичные меристемы
Вторичная боковая (латеральная) меристема представлена камбием и феллогеном и формируется из промеристем (прокамбия) или постоянных тканей путем их дедифференцировки. Клетки камбия делятся перегородками, параллельными поверхности органа (периклинально). Из клеток, отложенных камбием наружу, развиваются элементы вторичной флоэмы, а внутрь из отложенных камбием - вторичной ксилемы. Камбий, возникший из постоянных тканей путем дедифференцировки, называют добавочным. По строению и характеру своей деятельности он не отличается от камбия, возник- шего из промеристем. Феллоген формируется из постоянных тканей, расположенных в субэпидермальных слоях (под эпидермой). Делясь параллельно поверхности органа, феллоген отделяет наружу будущие клетки пробки (феллемы), а внутрь - клетки феллодермы. Таким образом, феллоген формирует вторичную покровную ткань - перидерму.
Боковые меристемы располагаются параллельно их поверхности, и обеспечивают рост осевых органов двудольных растений в толщину.
Раневые меристемы образуются при повреждении тканей и органов. Вокруг повреждения живые клетки дедифференцируются, начинают делиться и тем самым превращаются во вторичную меристему. Их задача - образовать плотную защитную ткань, состоящую из паренхимных клеток, - каллюс. Это ткань белого или желтоватого цвета, которая возникает при прививках, обеспечивая срастание привоя с подвоем и в основании черенков.
Покровные ткани
Первичной покровной тканью является эпидерма, которая состоит из нескольких типов клеток: собственно-эпидермальных, околоустьичных, замыкающих клеток устьица и трихом. Наружные стенки клеток эпидермы покрыты пленкой - кутикулой. У некоторых растений (агавы, кливии, лука) толстые стенки листьев имеют несколько кутинизированных слоев, чередующихся со слоями целлюлозы. Снаружи откладывается чистый кутин, образуя непрерывный слой кутикулы различной толщины. Далее вглубь следуют так называемые кутикулярные слои стенки, состоящие из целлюлозы, пектиновых веществ, радиально расположенных слоев воска и распределенного в беспорядке между ними слоев кутина. Самый внутренний слой клеточной стенки, прилегающий к полости клетки, не содержит кутина. Кутикула иногда вклинивается между боковыми стенками клеток (рис. 2.3). Она защищает клетки от механических повреждений, проникновения паразитов и поглощает ультрафиолетовые лучи.
Кутинизация рассматривается как приспособление для уменьшения испарения (транспирации) и вымывания веществ из клеток выпадающими осадками.
Пектиновые вещества и целлюлоза, входящие в клеточную стенку, могут подвергаться ослизнению с образованием слизей и камедей. Они представляют собой полимерные углеводы, родственные пектиновым веществам, и отличаются способностью к сильному набуха нию при соприкосновении с водой. Камеди в набухшем состоянии клейки и могут вытягиваться в нити, тогда как слизи сильно расплываются и в нити не вытягиваются. Пектиновые слизи встречаются у представителей следующих семейств: лилейных, крестоцветных, мальвовых, липовых, розоцветных, в отличие от целлюлозных слизей, встречающихся гораздо реже (например, у орхидных).
Латеральные (боковые) меристемы по происхождению могут быть первичными и вторичными. На поперечном срезе осевых органов они имеют вид колец. Примером первичной боковой меристемы являются прокамбий, перицикл. Прокамбий - меристема, из которой формируются первичные элементы сосудисто-волокнистых пучков (первичные флоэма и ксилема) и камбий - вторичная меристема. При этом клетки прокамбия непосредственно дифференцируются в клетки первичных проводящих тканей.
Латеральные (боковые) меристемы по происхождению могут быть первичными и вторичными. На поперечном срезе осевых органов они имеют вид колец. Примером первичной боковой меристемы являются прокамбий, перицикл. Прокамбий - меристема, из которой формируются первичные элементы сосудисто-волокнистых пучков (первичные флоэма и ксилема) и камбий - вторичная меристема. При этом клетки прокамбия непосредственно дифференцируются в клетки первичных проводящих тканей.
Латеральные (боковые) меристемы по происхождению могут быть первичными и вторичными. На поперечном срезе осевых органов они имеют вид колец. Примером первичной боковой меристемы являются прокамбий, перицикл. Прокамбий - меристема, из которой формируются первичные элементы сосудисто-волокнистых пучков (первичные флоэма и ксилема) и камбий - вторичная меристема. При этом клетки прокамбия непосредственно дифференцируются в клетки первичных проводящих тканей.
Латеральные (боковые) меристемы по происхождению могут быть первичными и вторичными. На поперечном срезе осевых органов они имеют вид колец. Примером первичной боковой меристемы являются прокамбий, перицикл. Прокамбий - меристема, из которой формируются первичные элементы сосудисто-волокнистых пучков (первичные флоэма и ксилема) и камбий - вторичная меристема. При этом клетки прокамбия непосредственно дифференцируются в клетки первичных проводящих тканей.
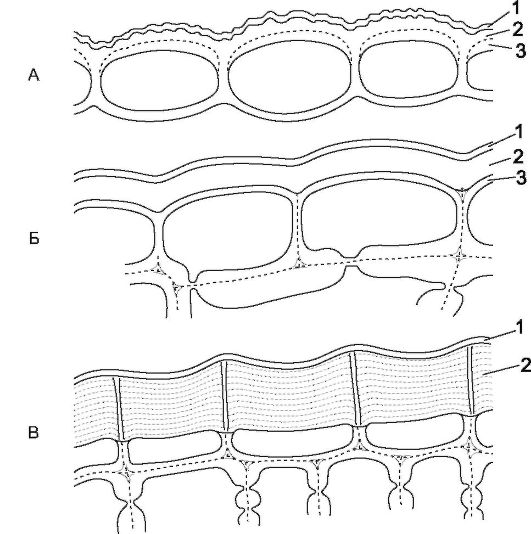
Рис. 2.3.Кутинизированная оболочка (поперечный срез): А - лист гвоздики перистой; Б - стебель Cereus triangularis (сем. кактусовых); В - стебель Kleinia neriifolia (сем. сложноцветных): 1 - кутикула; 2 - кутикулярные слои; 3 - целлюлозный слой наружной стенки клеток
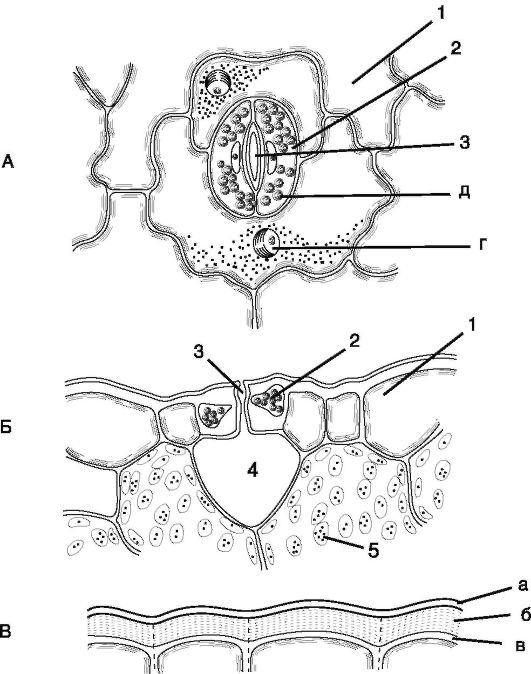
Рис. 2.4.Строение устьица: А, Б - кожица листа тимьяна (вид сверху и на поперечном срезе); В - кожица со стебля цереус (сем. кактусовых); 1 - собственно эпидермальные клетки; 2 - замыкающие клетки устьица; 3 - устьичная щель; 4 - воздухоносная полость; 5 - клетки хлорофиллоносной паренхимы; а - кутикула; б - кутикулярный слой - оболочка с суберином и воском; в - целлюлозный слой стенки; г - ядро с ядрышком; д - хлоропласты
Устьица представляют собой высокоспециализированные образования эпидермы, состоящие из 2 замыкающих клеток бобовидной формы и устьичной щели (своеобразного межклетника между ними). Имеются главным образом в листьях, но встречаются и на стебле (рис. 2.4).
Стенки замыкающих клеток утолщены неравномерно: клеточные стенки, направленные к щели (брюшные), значительно утолщены по сравнению с направленными от щели (спинные стенки). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью располагается крупная полость (межклетник), называемая дыхательной, она окружена клетками мезофилла листа.
Устьица обычно располагаются на нижней стороне листа. Однако у водных растений, обладающих плавающими листьями, они находятся только на верхней стороне листа. По форме клеток эпидермы и рас- положению устьиц можно отличить однодольное растение от двудольного (рис. 2.5).
Механизм работы устьиц обусловлен осмотическими свойствами клеток. При освещении поверхности листа солнцем в хлоропластах замыкающих клеток идет активный процесс фотосинтеза. Насыщение клеток продуктами фотосинтеза - сахарами - влечет за собой активное поступление в клетки ионов калия, вследствие чего концентрация клеточного сока в замыкающих клетках возрастает. Возникает разность концентрации клеточного сока околоустьичных и замыкающих клеток.
В силу осмотических свойств клеток вода из околоустьичных клеток поступает в замыкающие клетки, что ведет к увеличению их объема и резкому возрастанию тургора. Утолщение брюшных стенок замыкающих клеток, обращенных к устьичной щели, обеспечивает неравномерное растяжение клеточной стенки; замыкающие клетки приобретают выраженную бобовидную форму и происходит открытие устьичной щели.
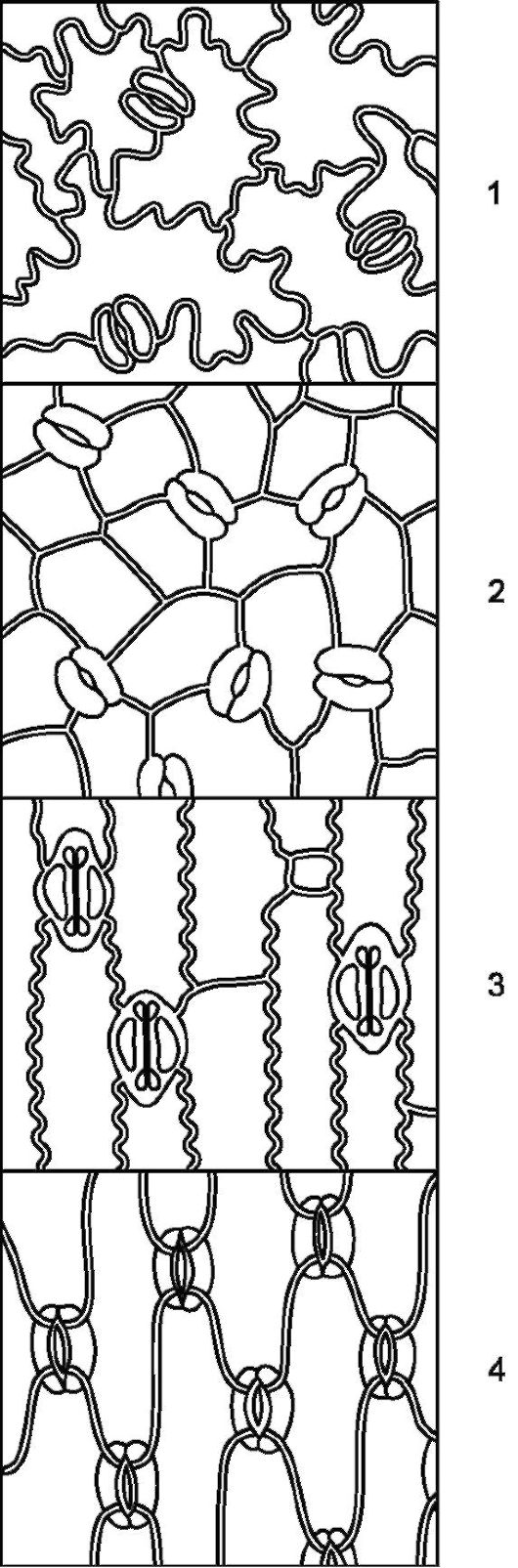
Рис. 2.5.Расположение устьиц на эпидерме (вид с поверхности): 1 - буквица; 2 - арбуз; 3 - кукуруза; 4 - ирис
При падении интенсивности фотосинтеза (например, вечером) снижается образование сахаров в замыкающих клетках. Приток ионов калия прекращается. Концентрация клеточного сока падает в замыкающих клетках по сравнению с таковой в околоустьичных. Вода путем осмоса уходит из замыкающих клеток, и тургор этих клеток понижается, что ведет к закрытию устьичной щели ночью.
Клетки эпидермы плотно сомкнуты между собой, благодаря чему она выполняет ряд функций:
1) препятствует проникновению внутрь растения болезнетворных организмов;
2) защищает внутренние ткани от механических повреждений;
3) осуществляет регуляцию газообмена и транспирации;
4) через нее выделяются вода, соли;
5) может функционировать как всасывающая ткань;
6) принимает участие в синтезе различных веществ, восприятии раздражений, движении листьев.
Часто в собственно-эпидермальных клетках содержатся амилопласты - классическая форма лейкопластов (см. рис. 1.5).
Трихомы - это различные по форме, строению и функциям выросты клеток эпидермы: волоски, чешуйки, щетинки и т.п. Они подразделяются на кроющие и железистые. Железистые трихомы в отличие от кроющих имеют секреторные клетки с секретом, являющимся продуктом выделения. Кроющие волоски образуют на растении шерстистый, войлочный или иной покров, отражая часть солнечных лу- чей, чем уменьшают транспирацию. Иногда волоски располагаются
только там, где расположены устьица, например на нижней стороне листа мать-и-мачехи. У некоторых растений живые волоски увеличи- вают общую испаряющую поверхность, что способствует ускорению транспирации.
Размеры трихом варьируют в значительных пределах. Наиболее длинные трихомы (до 5-6 см) покрывают семена хлопчатника. Кроющие трихомы имеют форму простых одноклеточных или многоклеточных, разветвленных или звездчатых волосков. Кроющие трихомы могут длительное время оставаться живыми или быстро отмирать и заполняться воздухом (рис. 2.6) (рис. 2, см. цв. вкл.).
От трихом, возникающих только при участии эпидермальных клеток, отличаются эмергенцы, в образовании которых участвуют и более глубоко расположенные ткани субэпидермального слоя (рис. 2.7).
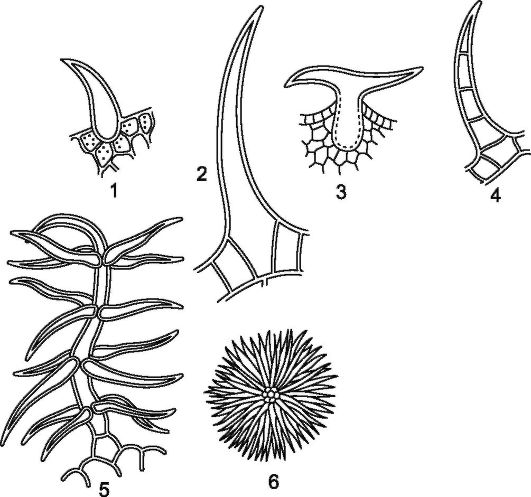
Рис. 2.6.Эпидермальные волоски: 1, 2 - подмаренника; 3 - хмеля; 4 - наперстянки; 5 - коровяка; 6 - лоха
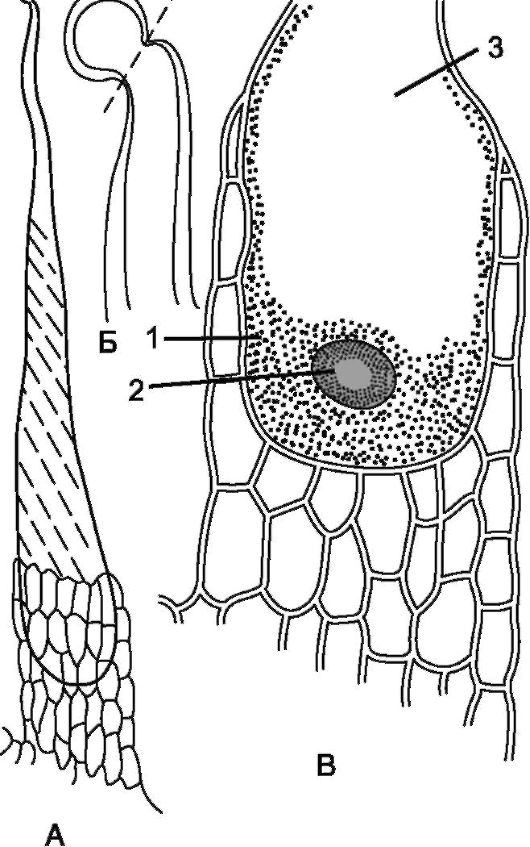
Рис. 2.7.Эмергенцы (у крапивы): А - общий вид; Б - окончания волоска (показана линия облома); В - основание волоска с цитоплазмой (1), ядром (2) и вакуолью (3)
Вторичная покровная ткань называется перидермой. Это сложная покровная ткань стеблей, корней и корневищ многолетних растений. Она сменяет эпидерму осевых органов, которая постепенно отмирает и слущивается. Перидерма образуется из феллогена. Последний закладывается в эпидерме, субэпидермальном слое и даже в глубоких слоях осевых органов. Клетки феллогена делятся и наружу откладывают клетки пробки, а внутрь - живые паренхимные клетки феллодермы. В клетках феллодермы стеблей содержатся хлоропласты.
Пробка состоит из мертвых клеток, у которых клеточная стенка пропитана жироподобным веществом - суберином. Клетки располагаются ровными рядами, имеют прямоугольную форму (на поперечном срезе), плотно прилегая друг к другу и формируя многослойный футляр. Пробка охраняет внутренние живые ткани от потери влаги, резких температурных колебаний и проникновения микроорганизмов. Живые ткани, лежащие под пробкой, нуждаются в газообмене и удалении избытка влаги. Поэтому под устьицем за счет деления субэпидермальных слоев, а в дальнейшем и феллогена откладываются живые, рыхло расположенные, со множеством межклетников, паренхимные клетки, называемые выполняющей тканью, которая разрывает эпидерму и создает возможность газообмена и транспирации с внешней средой. Это структурное образование называется чечевичкой (рис. 2.8).
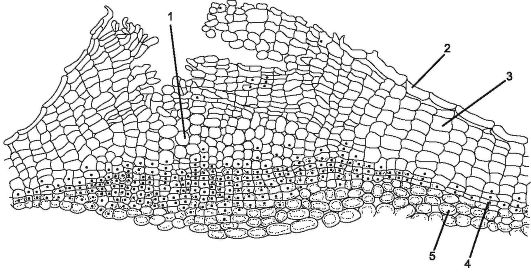
Рис. 2.8.Строение перидермы с чечевичкой: 1 - выполняющая ткань чечевички; 2 - остатки эпидермы; 3 - пробка (феллема); 4 - феллоген; 5 - феллодерма
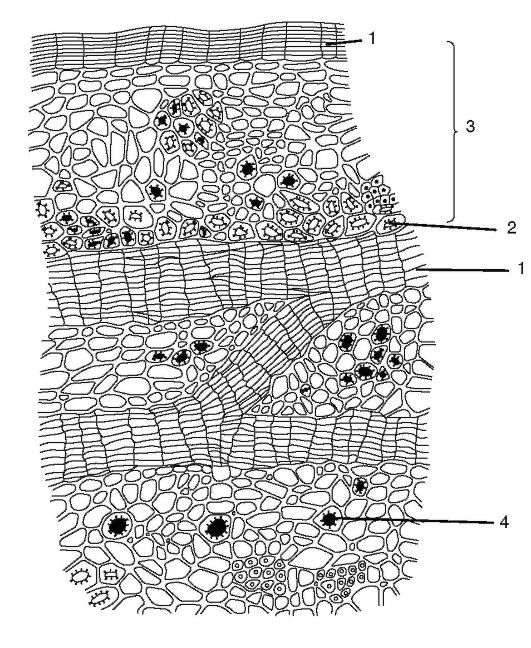
Рис. 2.9.Корка дуба: 1 - слои пробки; 2 - волокна; 3 - остатки первичной коры; 4 - друзы оксалата кальция
Чечевички, имеющие вид небольших бугорков, отчетливо видны на поверхности побегов деревьев и кустарников. На стволах березы их остатки наблюдаются в виде характерных поперечных черных полосок, у осины они принимают форму ромбов.
Корка (ритидом) является третичной покровной тканью,которая образуется у многолетних растений в корне, стебле, корневище. Каждый год в более глубоких слоях закладывается новый слой феллогена и образуется перидерма. Наружный слой перидермы - пробка - изолирует все вышележащие ткани, в результате чего они отмирают. Таким образом, совокупность многочисленных перидерм с отмершими между ними тканями и является коркой (рис. 2.9).
Основные ткани
Основные ткани составляют большую часть всех органов растений. Они заполняют промежутки между проводящими и механиче- скими тканями и присутствуют во всех вегетативных и генеративных органах. Эти ткани образуются за счет дифференцировки апикальных меристем и состоят из живых паренхиматозных клеток, разнообразных по строению и функциям. Различают ассимиляционную, запасающую, воздухо- и водоносную паренхимы.
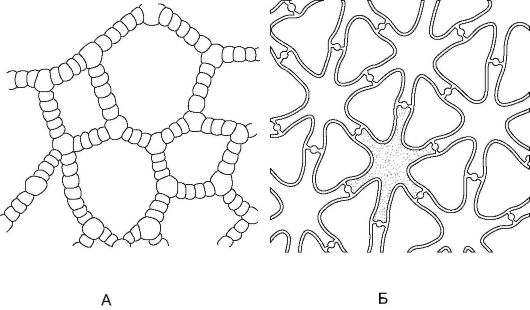
Рис. 2.10.Аэренхима в черешке листа кувшинки (А) и в стебле ситника развесистого (Б)
В ассимиляционной, или хлорофиллоносной, паренхиме осуществляется фотосинтез. Эта ткань встречается в надземных органах растений (листьях, молодых зеленых стеблях).
Запасающая паренхима преобладает в стебле, корне, корневище. В клетках этой ткани откладываются запасающие вещества - белки, жиры, углеводы.
Воздухоносная паренхима, или аэренхима, состоит из воздухоносных полостей (межклетников), представляющих собой резервуары для запаса газообразных веществ. Эти полости окружены клетками основной паренхимы (хлорофиллоносной или запасающей). Аэренхима хорошо развита у водных растений в различных органах и может встречаться у сухопутных видов; главное ее назначение - участие в газообмене, а также в обеспечении плавучести растений (рис. 2.10).
Клетки водоносной паренхимы содержат в вакуолях слизистые вещества, способствующие удержанию влаги. Преимущественно эти клетки бывают у суккулентов (кактусы, алоэ, агава).
Механические ткани
Механические ткани - это опорные (арматурные) ткани, образующие скелет растения и обеспечивающие его прочность, вследствие чего растение способно противостоять нагрузкам на растяжение, сжатие и изгиб. Различают механические ткани с равномерно и неравномерно утолщенными клеточными стенками.
Колленхима.Это первичная по происхождению ткань, клетки которой имеют неравномерно утолщенные клеточные стенки. Различают уголковую, пластинчатую и рьхлую колленхиму (рис. 2.11). Клеточная стенка колленхимы состоит из целлюлозы, гемицеллюлозы и пектиновых веществ. Клетки колленхимы являются хлорофиллоносными, поэтому в подземных органах не встречаются. Эволюционно колленхима возникла из паренхимы. Формируется она из основной меристемы и находится под эпидермой или на расстоянии одного (или нескольких) слоев от нее.
Клетка уголковой колленхимы имеет форму вытянутого шестиугольного многогранника, у которого утолщение целлюлозной оболочки идет вдоль ребер, а на поперечном срезе утолщения клеточной стенки заметны по углам этого многогранника. Уголковая колленхима встречается по периферии стеблей двудольных растений (в основном травянистых), в черешках листьев и по обеим сторонам крупных
жилок листа. Колленхима не препятствует росту органа, в котором она расположена, в длину.
Клетка пластинчатой колленхимы имеет форму параллелепипеда, у которого утолщаются только тангентальные стенки (параллельные поверхности стебля). Пластинчатая колленхима встречается, как правило, в стеблях древесных растений, но может быть и в травянистых (у стебля подсолнечника). Клетки уголковой и пластинчатой колленхимы расположены плотно друг к другу, не образуя межклетни- ков. Рыхлая колленхима имеет межклетники, а утолщенные клеточные стенки направлены в сторону этих межклетников.
Склеренхимные волокна и склереиды.Механическая ткань, состоящая из клеток с одревесневшими и равномерно утолщенными клеточными стенками, называется склеренхимой. Ее клеточные стенки одревесневают, т. е. пропитываются лигнином, а ядро и цитоплазма клетки разрушаются. Склеренхим ные волокна образуют ткань, состоящую из клеток вытянутой формы с заостренными концами и поровыми каналами в клеточных стенках. Эти клетки плотно примыкают друг к другу, и их стенки обладают высокой прочностью. На поперечном срезе клетки многогранны. По происхождению склеренхима может быть первичной, возникающей из прокамбия или перицикла, и вторичной - из клеток камбия.

Рис. 2.11.Колленхима: А - уголковая колленхима; Б - поперечный разрез через пластинчатую колленхиму; В - рыхлая колленхима с межклетниками; Г - объемное изображение уголковой колленхимы: 1 - постенный слой цитоплазмы; 2 - ядро; 3 - вакуоль; 4 - утолщенная оболочка
Если склеренхимные волокна встречаются в древесине (ксилеме), то они называются древесинными волокнами (либриформ) (рис. 2.12). Являясь механической частью ксилемы, древесинные волокна защи- щают сосуды от давления других тканей. Если склеренхимные волокна встречаются в лубе (флоэме), они называются лубяными волокнами (камбиформ) (см. рис. 2.12). Лубяные волокна могут быть и неодревесневшими, обладая при этом большой прочностью и эластичностью, что находит применение в текстильной промышленности (например, волокна льна). Если волокна возникают на месте перицикла, они называются перициклическими, а если в коре - кóровыми.
Склеренхимные клетки, не обладающие формой волокна, называются склереидами. Склереиды обычно возникают из клеток основной паренхимы в результате утолщения и лигнификации их клеточных стенок. Они бывают различной формы и встречаются во многих органах растения. Склереиды более или менее изодиаметричной формы (с одинаковым диаметром клетки) называются брахисклереидами (рис. 2.13) или каменистыми клетками (в плодах груши). Склереиды в форме берцовой кости с расширением на обоих концах клетки - остеосклереиды - встречаются в листьях чая. Склереиды, форма которых напоминает звезду, называются астросклереидами (например, в листьях камелии). Удлиненные палочковидные клетки склереид на- ходятся, например, в семенах бобовых (макросклереиды).
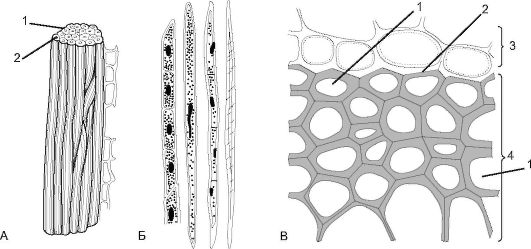
Рис. 2.12.Склеренхима: А - лубяные волокна: 1 - утолщенная оболочка; 2 - полость клетки; Б - древесинные волокна (либриформ); В - на поперечном срезе стебля тыквы: 1 - полость клетки склеренхимы; 2 - утолщения оболочки; 3 - паренхимные клетки; 4 - склеренхима
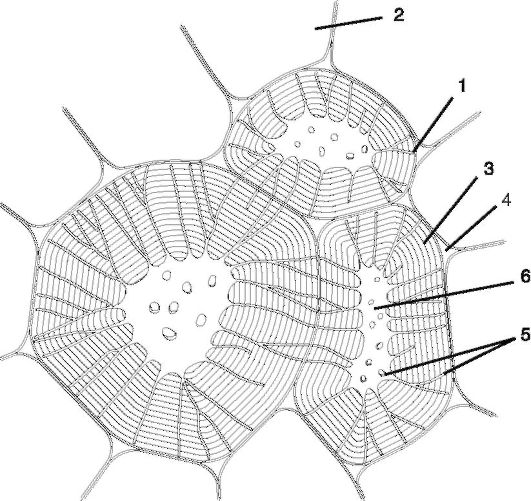
Рис. 2.13.Каменистые клетки из околоплодника груши:
1 - первичная оболочка;
2 - тонкостенные клетки; 3 - слои вторичной оболочки; 4 - межклетники; 5 - поровые каналы в плане и в разрезе; 6 - полость клетки
Проводящие ткани
Проводящие ткани обеспечивают восходящий и нисходящий ток растения. Восходящий ток - это ток минеральных солей, растворенных в воде, идущих от корней по стеблю к листьям. Восходящий ток осуществляется по сосудам и трахеидам ксилемы(древесины). Нисходящий ток - это ток органических веществ, направляющийся от листьев к корням по ситовидным элементам флоэмы(луба).
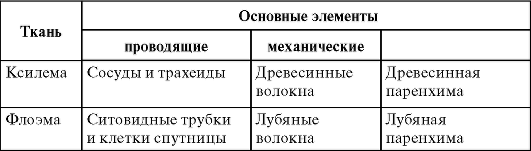
Ксилема и флоэма - это сложные ткани, состоящие из 3 основных элементов (табл. 2.2).
Проводящие элементы ксилемы.Наиболее древними проводящими элементами ксилемы являются трахеиды - это вытянутые клетки с заостренными концами. Трахеиды имеют одревесневшую клеточную стенку. По характеру утолщения оболочек, размерам и расположению в них участков первичных оболочек различают 4 типа трахеид: кольчатые, спиральные, пористые и лестничные (рис. 2.14). К наиболее древним пористым трахеидам относят лестничные трахеиды.
Таблица 2.2.Основные элементы ксилемы и флоэмы
Сосуды (или трахеи) представляют собой однорядный продольный тяж клеток, называемых члениками. В филогенезе членики трахеи произошли из трахеид. Благодаря перфорациям между члениками вдоль всего сосуда свободно осуществляется ток жидкости. Утолщения клеточных оболочек у сосудов бывают кольчатыми, спиральными, лестничными, сетчатыми и пористыми (точечными).
Проводящие элементы флоэмыу архегониальных растений, кроме мхов, представлены си- товидными клетками. На их продольных стенках имеются сквозные отверстия, напоминающие сито, а потому называемые ситовидными полями. У покрытосеменных растений в процессе эволюции сформировался 2-й тип проводящих элементов - ситовидные трубки, представляющие собой продольный тяж клеток, называемых члениками. Конечные стенки члеников превращены в ситовидные пластинки: простые (имеют по одному ситовидному полю) и сложные (имеют несколько ситовидных полей). Ситовидные поля могут встречаться и на боковых стенках члеников (у березы). В ситовидном элементе разрушается тонопласт («оболочка» вакуоли), и вакуолярный сок смешивается с гиалоплазмой. Ядро, как правило, при этом разрушается. Каждому членику ситовидной трубки сопутствуют (одна или несколько) специализированных паренхимных клеток - (клетки-спутницы), выполняющие вспомогательную роль в транспорте органических веществ (рис. 2.15). Передвижение по ситовидным трубкам - физиологический процесс, идущий под действием ферментов выделяемых клетками-спутницами.
Сосудисто-волокнистые пучки.Флоэма и ксилема образуют сосудисто-волокнистые пучки, которые располагаются в центральном осевом цилиндре и бывают открытыми и закрытыми.
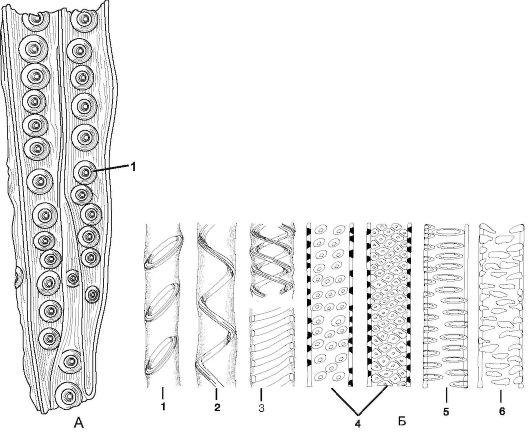
Рис. 2.14.Типы трахеид и сосудов: А - трахеиды древесины сосны: 1 - окаймленная пора. Б - типы утолщения и поровости боковых стенок у сосудов: 1 - кольчатое; 2, 3 - спиральные; 4 - сетчатое; 5 - лестничное; 6 – супротивное
Закрытые пучки состоят из ксилемы и флоэмы, между которыми отсутствует камбий и, таким образом, не происходит образования новых элементов флоэмы и ксилемы. Закрытые сосудисто-волокнистые пучки встречаются в стеблях и корневищах однодольных растений.
Открытые пучки имеют камбий между флоэмой и ксилемой. В результате деятельности камбия пучок разрастается и происходит утолщение органа. Открытые сосудисто-волокнистые пучки встречаются во всех осевых органах двудольных и голосеменных растений.
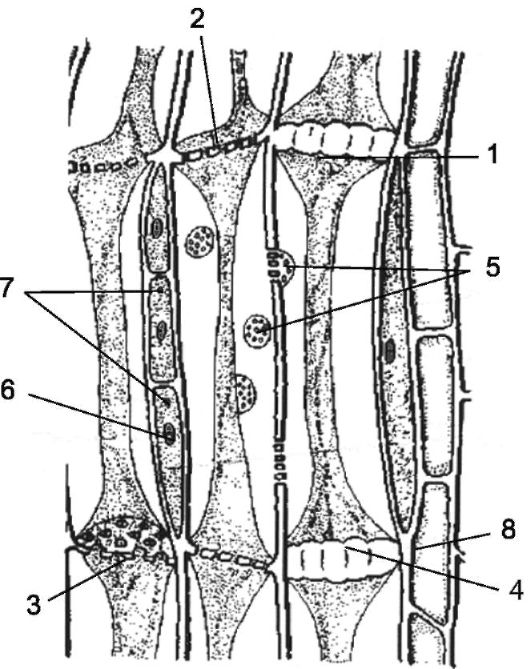
Рис. 2.15.Продольный срез флоэмы: 1 - членик ситовидной трубки с тяжами содержимого; 2 - ситовидные пластинки; 3 - ситовидные отверстия; 4 - мозолистое тело; - ситовидные поля на боковых стенках; 6 - ядро; 7 - сопровождающие клетки;8 - флоэмная паренхима
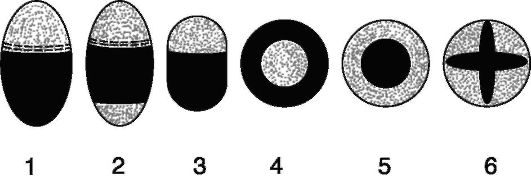
Рис. 2.16.Схема разных типов проводящих сосудисто-волокнистых пучков (поперечные срезы): 1 - открытый коллатеральный; 2 - биколлатеральный; 3 - закрытый коллатеральный; 4, 5 - концентрические (4 - центрофлоэмный, 5 - центроксилемный); 6 - радиальный. Ксилема зачернена, флоэма показана точками, камбий - штрихами
В зависимости от взаимного расположения флоэмы и ксилемы различают 4 типа сосудисто-волокнистых пучков (рис. 2.16).
Чаще всего флоэма лежит по одну сторону от ксилемы, т.е. она примыкает к ксилеме и обращена к периферии органа. Такие пучки называют коллатеральными. Они могут быть как закрытыми, так и открытыми. Закрытые коллатеральные пучки встречаются у стеблей и корневищ однодольных растений, а открытые - у стеблей, корневищ и корней двудольных растений.
В биколлатеральных пучках флоэма примыкает к ксилеме с 2 сторон. Мощный участок флоэмы обращен к периферии органа и отделен от ксилемы слоем камбия. Внутренняя флоэма развита слабо и примыкает к ксилеме без камбия. Биколлатеральные пучки встречаются у стеблей, корней и корневищ двудольных растений.
Радиальные пучки составлены ксилемой, которая располагается по радиусам. Между лучами ксилемы находятся участки флоэмы. Эти пучки не имеют камбия и встречаются только у корней в первичном строении.
Концентрические пучки складываются из ксилемы, окружающей флоэму, - центрофлоэмные пучки (у большинства корневищ однодольных), а центроксилемные - когда флоэма окружает ксилему (у корневищ папоротников).
Дата добавления: 2016-05-30; просмотров: 10147;
Поиск по сайту
Узнать еще
- Артериосклероз. Атеросклероз. Артериальная гипертензия: гипертоническая болезнь и вторичные артериальные гипертензии. Ишемические болезни сердца (ИБС)
- ВТОРИЧНЫЕ ВИДЫ ХОЗЯЙСТВЕННОЙ ДЕЯТЕЛЬНОСТИ
- ВТОРИЧНЫЕ ЗЛОКАЧЕСТВЕННЫЕ ОПУХОЛИ КОСТЕЙ
- Вторичные источники питания
- ВТОРИЧНЫЕ МОРФОЛОГИЧЕСКИЕ ЭЛЕМЕНТЫ
- Вторичные морфологические элементы сыпи
- ВТОРИЧНЫЕ НАУЧНЫЕ ТЕКСТЫ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
