Геномная организация генов, кодирующих иммуноглобулины
Огромное разнообразие молекул иммуноглобулинов в пределах каждого класса обусловлено:
Ø во-первых, большим количеством генов, кодирующих разные участки (константные и вариабельные) тяжелых и легких цепей иммуноглобулинов,
Ø во-вторых, определенными изменениями,происходящими в геноме незрелых В-лимфоцитов в процессе их деления и созревания.
Для человека и млекопитающих животных известны 3 группы сцепления генов, кодирующих иммуноглобулины. Эти группы сцепления расположены в разных парах хромосом. В частности, существует группа сцепления генов, кодирующих тяжелые цепи, и две разные группы сцепления, кодирующие c- и l-типы легких цепей. Каждая группа сцепления, в свою очередь, представлена большим количеством генов, одни из которых кодируют вариабельную область цепи (V-гены), а другие – константную область (С-гены). Причем эти V- и С-гены какой-то определенной группы сцепления изначально (в незрелом В-лимфоците) находятся на значительном расстоянии друг от друга. Подобная нативная локализация генов, кодирующих цепи иммуноглобулинов, определяется как состояние зародышевой линии (англ. germline). Однако по мере созревания В-лимфоцитов и соответственно перехода их от некоммитированных предшественников к коммитированым формам, проявляющим определенную специфичность, происходит реорганизация генома, так что пространственно удаленные гены оказываются расположенными в непосредственной близости друг от друга, образуя единый информационный участок. Такая реорганизация генома получила название соматической рекомбинации и осуществляется путем делеции (вырезания) определенных генов в группе сцепления, в результате чего далеко отстоящие гены оказываются расположенными друг возле друга. Соматическая рекомбинация затрагивает только лишь определенные соматические клетки (в нашем случае развивающиеся В-лимфоциты), не происходит в половых клетках, а, следовательно, не наследуется.
Группа сцепления, ответственная за синтез легкой цепи c-типа, включает три группы генов:
ü 250 генов, кодирующих область вариабельного домена c-цепи (Vc-гены), состоящую из 94-95 аминокислотных остатков. Причем каждому Vc-гену в полинуклеотидной последовательности обязательно предшествует т.н. лидерный участок (L-ген), кодирующий т.н. лидерный пептид, состоящий из 20-25 аминокислотных остатков, обладающий гидрофобностью и облегчающий прохождение молекулы иммуноглобулина через мембрану гранулярного эндоплазматического ретикулума. Лидерный пептид присущ только незрелой молекуле иммуноглобулина, после прохождения этой незрелой молекулы через мембрану эндоплазматического ретикулума он отщепляется. Каждому из 250 Vc-генов предшествует один и тот же L-ген.
ü 5 J-мини-генов (от англ. "joining"), кодирующих аминокислотную последовательностьиз 12-14 аминокислот, достраивающую вариабельный домен. Один из этих 5 J-мини-генов является неактивным и в связи с этим называется псевдогеном
ü 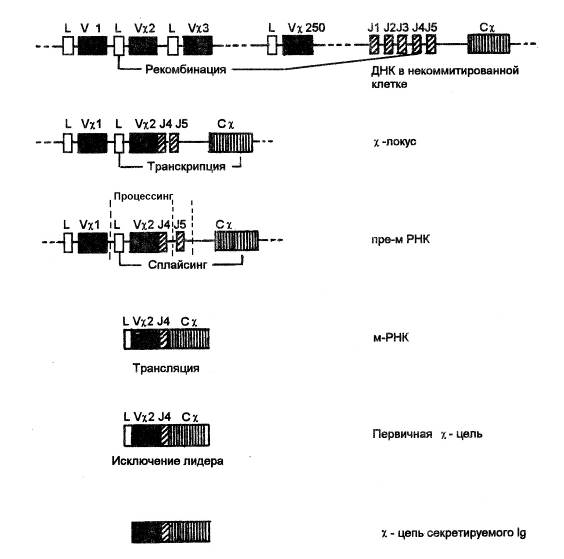 один ген, кодирующий константную область c-цепи (Сc-ген).
один ген, кодирующий константную область c-цепи (Сc-ген).
Рис. Рекомбинация генов, кодирующих c-цепь иммуноглобулинов. В некоммитированном В-лимфоците гены, кодирующие c-цепь иммуноглобулинов, отдалены друг от друга в ДНК на значительное расстояние. Процесс рекомбинации начинается с объединения одного из 250 V-генов (в данном случае Vc2) с одним из 5 J-мини-генов (в данном случае J4). В результате формируется c-локус, контролирующий незрелую пре-мРНК ядра. При созревании такой РНК происходит процессинг пре-мРНК, в ходе которого вырезаются ее участки, комплементарные областям ДНК, расположенным между кодирующими генами c-локуса. Затем участки РНК, комплементарные LVc2J4 и Сc -генам объединяются в результате сплайсинга, что приводит к образованию зрелой мРНК, транслирующей первичную структуру c-цепи. При созревании c-цепи лидерная полипептидная последовательность удаляется, что и приводит к формированию окончательной c-цепи.
Процесс рекомбинации начинается с объединения одного из 250 Vc-генов с одним из 5 J-мини-генов (за счет делеции участков ДНК между этими генами) и осуществляется случайным образом. Место рекомбинации V-J не является жестко фиксированным: объединение между определенными V- и J-геном может происходить как между собственно их основаниями, так и между областями, соседствующими с этими V- и J-генами. Подобные "ошибки" вносят дополнительную изменчивость в третью гипервариабельную область вариабельного домена легкой цепи. Образовавшийся вследствие делеций информационный участок VicJic далее путем делеций объединяется с Сc-геном, в результате чего формируется c-локус, состоящий из трех экзонов (кодирующих участков):
ü L-гена, кодирующего лидерный пептид
ü рекомбинантного VicJic-гена, кодирующего вариабельный домен легкой χ-цепи
ü Сc-гена,кодирующего константный домен легкой χ-цепи.
Определенная структура возникающего вследствие рекомбинации генома c-локуса и будет обуславливать специфику вариабельного домена синтезируемой в таком В-лимфоците c-цепи. Рекомбинантная ДНК в коммитированном на определенный антиген В-лимфоците обеспечивает образование первичного транскрипта – пре-мРНК ядра, включающей полинуклеотидные последовательности, считанные как с трех экзонов c-локуса (L-гена, рекомбинантного VicJic-генаи Сc-гена), так и с некодирующих участков (интронов), находящихся между L-геномирекомбинантным VicJic-геном,а также междуVicJic-геноми Сc-геном.Образовавшаяся пре-мРНК ядрадалее (еще до выходя из ядра) подвергается созреванию, включающему
ü процессинг пре-мРНК ядра, в ходе которого из нее вырезаются интронные участки
ü сплайсинг экзонных участков пре-мРНК,в результате которого образуется зрелая мРНК иммуноглобулина, покидающая ядро и транслирующаяся на рибосомах, связанных с мембраной эндоплазматического ретикулума.
Группа сцепления генов, кодирующих легкую цепь l-типа, представлена генами, локализованными в двух локусах, каждый из которых содержит:
ü один L-ген (обозначаемые L1-ген в первом локусе и L2-ген во втором локусе)
ü один Vl-ген (обозначаемые Vl1-ген в первом локусе и Vl2-ген во втором локусе)
ü два Jl-мини-гена (обозначаемые Jl1- и Jl2 –гены в первом локусе и Jl3- и Jl4 –гены во втором локусе)
ü два Сl-гена (обозначаемые Сl1- и Сl2 –гены в первом локусе и Сl3- и Сl4 –гены во втором локусе).
Гены Jl4 и Сl4, заложенные во втором локусе, являются псевдогенами (функционально не активны).
Несмотря на несколько иную организацию группы сцепления, кодирующей легкую цепь l-типа,процессы рекомбинации ДНК при созревании В-лимфоцитов и последующего созревания пре-мРНК ядра происходят аналогично таковым при синтезе легкой χ-цепи. При этом возможны следующие варианты рекомбинантной ДНК и соответственно разные типы антител, кодируемых ими:
ü L1 Vl1 Jl1 Сl1
ü L1 Vl1 Jl2 Сl2
ü L2 Vl2 Jl3 Сl3
Образование еще одного теоретически возможного варианта – L2 Vl2 Jl4 Сl4 – практически никогда не осуществляется, поскольку гены Jl4 и Сl4 являются не активными.
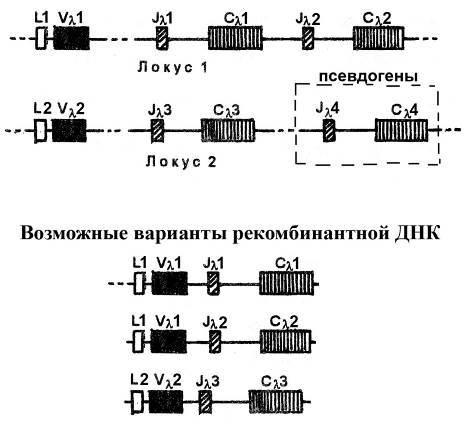
Рис. Геномная организация генов, кодирующих легкую цепь иммуноглобулинов l-типа. l-цепь кодируется двумя локусами, каждый из которых включает один L-ген, один Vl-ген, два Jl-мини-гена и два Сl-гена. В локусе 2 гены Jl4 и Сl4 являются функционально неактивными (псевдогенами).
Тяжелые цепиразных изотипов (a, g, m, d, e) кодируются следующими группами генов:
ü группой генов, кодирующих вариабельный домен тяжелой цепи (V-домен).Эта группа включает следующие гены:
· 500 V-генов, каждому из которых предшествует свой L-ген, кодирующий лидерный пептид
· 15 D-мини-генов
· 4 J-мини-гена
ü группой генов, кодирующих константный участок тяжелой цепии представленных:
· 10 C-генами, определяющими изотип тяжелой цепи (a или g или m или d или e), а также особенности тяжелых цепей изотипа g (g1, g2а, g2b, g3)
При созревании В-лимфоцитов в группе сцепления генов, кодирующих тяжелые цепи иммуноглобулинов, происходит два рекомбинационных события:
Ø объединение определенного (любого из 15-ти) D-мини-гена с определенным (каким-то из 4-х) J-мини-геном в результате делеции участка ДНК между этими генами, в результате чего возникает определенныйDiJi-участок
Ø объединение сформированного DiJi-участка с одним из 500 V-генов (каждому V-гену, в свою очередь, предшествует свой L-ген). Такое объединение также происходит путем делеции участка ДНК между LiVi-участкоми DiJi-участком. Таким образом, возникает LiViDiJi-область,кодирующая синтез определенного вариабельного домена тяжелой цепи.
После рекомбинации ДНК образовавшаяся LiViDiJi-область, кодирующая вариабельный домен тяжелой цепи,функционально объединяется сСm-геном,кодирующим константную область m-типа(т.е. константную область иммуноглобулина класса М).Такое функциональное объединение достигается благодаря тому, чтопродукт транскрипции LiViDiJi-областипутемсплайсингасоединяется спродуктом транскрипции Сm-гена,в результате чего формируется зрелая мРНК, которая поступает из ядра в цитоплазму и транслируется на рибосомах эндоплазматического ретикулума. Следовательно, на начальном этапе дифференцировки В-лимфоцитов они синтезируют преимущественно иммуноглобулин М. По мере же функционального созревания В-лимфоцитов наблюдается переключение синтеза иммуноглобулина М на иммуноглобулины других классов (G, A, D или Е); при этом сохраняется исходная специфичность вариабельного домена (кодируемого рекомбинантным участком LiViDiJi-областиДНК),но при этомLiViDiJi-областьфункционально взаимодействует с С-генами, кодирующими константные области других изотипов тяжелых цепей(Сa или Сe и т.д.).
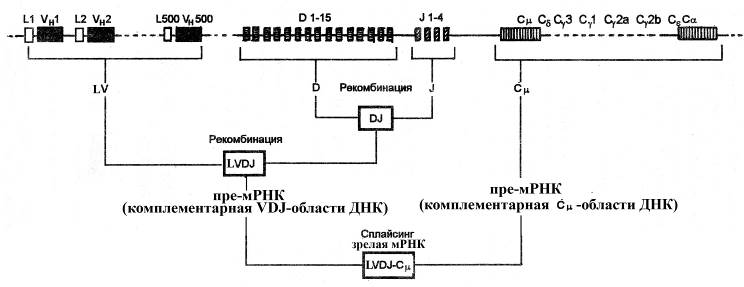
Рис. Рекомбинация генов, кодирующих тяжелые цепи иммуноглобулинов. Тяжелые цепи иммуноглобулинов кодируются четырьмя типами генов: V (каждому из которых предшествует L-ген), D, J и С. Первое рекомбинационное событие – объединение одного из 15 D-мини-генов с одним из 4 J-мини-геном. Второе рекомбинационное событие – объединение одного из 500 V-генов (каждому V-гену предшествует L-ген) с определенным DJ-участком. Следующий этап – функциональное объединение LVDJ-участка с Сm-геном. Особенность реорганизации участка генома, кодирующего тяжелые цепи В-лимфоцитов, состоит в переключении контроля синтеза иммуноглобулинов одного класса на другой при сохранении контроля специфичности реорганизованного LVDJ-локуса.
Таким образом, в процессе развития отдельных клонов В-лимфоцитов возможны две формы реорганизации участка генома, кодирующего тяжелые цепи иммуноглобулинов:
Ø в частности, на раннем этапе развития В-лимфоцитов определенные рекомбинантные LiViDiJi-области,разные в разных клонах В-лимфоцитов, объединяются с одним и тем жеС-геном (а именно, Сm-геном), что и определяет различия между разными клонами В-лимфоцитов
Ø в дальнейшем в каждом конкретном клоне В-лимфоцитов одна и та же LiViDiJi-область функционально объединяется(т.е. образует информационный участок) с разными С-генами, что обеспечивает внутриклеточное переключение синтеза иммуноглобулина одной и той же специфичности с одного класса на другой.
Изучение хромосомной организации иммуноглобулиновых генов и этапов их реорганизации в процессе развития В-лимфоцитов позволило определить причину вариабельности антител. Основу этой вариабельности составляет случайное объединение отдельных генных сегментов в результате рекомбинации ДНК: V, D, J-генов для тяжелых цепей и V, J-генов для легких цепей иммуноглобулинов. Если число V-генов тяжелых цепей равно 500 (не исключено, что их больше), D-сегментов – 15 и J-сегментов – 4, то число возможных вариантов V-доменов при использовании только этих показателей составит величину 30000 (500·15·4). Учитывая возможные нарушения, связанные с включением пограничных нуклеотидов справа и слева от D- и J-сегментов, число возможных вариантов V-доменов составит 120000 (30000·4). Расчет для V-доменов легкой цепи χ-типа даст возможное число вариантов этих доменов 2000 (250Vχ-генов·4J-мини-гена·2 (множитель, отражающий результаты ошибок при считывании пограничных нуклеотидов)). Поскольку молекулы иммуноглобулинов строятся путем случайного сочетания тяжелых и легких цепей, общее число вариантов антигенсвязывающих центров антител, образованных вариабельными доменами легкой и тяжелой цепей и определяющих специфичность иммуноглобулинов, окажется равным 2,4·108 (120000 вариантов V-доменов тяжелых цепей·2000 вариантов V-доменов легких цепей).
Таким образом, благодаря многочисленности генов, кодирующих определенные участки полипептидных цепей иммуноглобулинов, и случайной их комбинации друг с другом в процессе созревания В-лимфоцитов обеспечивается возможность создания огромного разнообразия антигенсвязывающих участков антител (до 2,4·108 различных вариантов вариабельных доменов антител), что обеспечивает надежную защиту организма от самых разнообразных антигенов.
Дата добавления: 2016-07-18; просмотров: 2808;
Поиск по сайту
Узнать еще
- II. Организация дезинфекционных и стерилизационных мероприятий в организациях, осуществляющих медицинскую деятельность
- II. Функционально-структурная организация и программное обеспечение персонального компьютера
- XXII. ОРГАНИЗАЦИЯ РАБОТ ПО ОБЕСПЫЛИВАНИЮ РУДНИЧНОГО ВОЗДУХА
- Автобусные маршруты и их организация
- Анатомическая организация мозга
- Базальные ганглии. Морфофункциональная организация. Функции
- Безопасная организация земляных работ
- Безопасная организация работ по очистке полосы отвода земли для строительства

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
