Контролируемых генами Х- и У-хромосом человека.
1. Сцепленное наследование признаков. Хромосомы как группы сцепления генов.
Менделевский закон независимого наследования применим лишь к тем случаям, когда гены, определяющие исследуемые признаки, лежат в разных хромосомах. Гораздо чаще мы сталкиваемся с явлением наследования нескольких признаков, гены которых лежат в одной и той же хромосоме.
У человека 23 пары хромосом, а генов более 20.000, следовательно, в одной и той же хромосоме находятся сотни генов. Гены, лежащие в одной хромосоме, образуют группу сцепления. Число групп сцепления равно числу хромосом в гаплоидном наборе. Хромосомы человека образуют 23 группы сцепления у женщин и 24 группы у мужчин (Y-хромосома образует отдельную группу сцепления). При мейозе гены данной группы сцепления попадают в одну гамету. Значит, наследоваться они будут одним организмом.
Сцепленное наследование изучал Т. Морган и его сотрудники. Т. Морган работал с дрозофилами – плодовыми мушками, которые быстро размножаются и неприхотливы к условиям содержания. Скрещивались гомозиготные мухи с серым телом (А) и длинными крыльями (В), с мухами, имеющими чёрное тело (а) и короткие крылья (в)
 Р
Р  ААВВ х
ААВВ х  аавв
аавв
 Г
Г
F1 АаВв
Все гибриды первого поколения имели серое тело и длинные крылья. Однако при скрещивании гибридов первого поколения Т. Морган не наблюдал явления независимого наследования признаков, т.е. у него не получалось расщепления в отношении 9 : 3 : 3 : 1 (как в опытах Г. Менделя).
Для того, чтобы узнать, какие гаметы образуют гибриды первого поколения Т. Морган провёл анализирующее скрещивание. Сначала он скрестил гетерозиготного самца и гомозиготную рецессивную самку. В потомстве ожидалось получить мух серых длиннокрылых, серых короткокрылых, чёрных длиннокрылых и чёрных короткокрылых, каждых по 25%.
Р 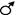 АаВв x аавв
АаВв x аавв
 |
Г
F1 АаВв, Аавв, ааВв, аавв
25% 25% 2 5% 25%
Такое расщепление должно получиться согласно законам Г.Менделя. Однако при анализе гибридов Т.Морган обнаружил серых мух с длинными крыльями (50%) и чёрных мух с короткими крыльями (50%). Для объяснения этого явления Т. Морган предположил, что гены, контролирующие серую окраску и длинные крылья, лежат в одной и той же хромосоме. Аллельные им гены, контролирующие чёрное тело и короткие крылья, лежат в гомологичной хромосоме.
Поэтому аллели (А) и (В) попадают в одну гамету, а аллели (а) и (в) в другую гамету, т.е. образуется 2 типа гамет, а не 4 как при независимом наследовании. Тогда и потомков во втором поколении будет 2 типа, а не 4. Следовательно, гены (А) и (В), а также (а) и (в) наследуются совместно, или сцеплено. Так как потомки, сочетающие признаки обоих родителей (серое тело, короткие крылья или чёрное тело, длинные крылья) отсутствуют, такое сцепление называется полным сцеплением.
2. Нарушение сцепления генов. Кроссинговер, его биологическая роль.
Когда Т. Морган скрестил гетерозиготную самку и гомозиготного рецессивного самца, в их потомстве наблюдалось расщепление. В процентном выражении это выглядело так: 41,5% серых длиннокрылых мух; 41,5% чёрных короткокрылых мух; 8,5% серых короткокрылых мух; 8,5% чёрных длиннокрылых мух.
Р АаВв х 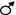 аавв
аавв
 Г
Г
F1 АаВв, Аавв, ааВв, аавв
с.д. с.к. ч.д. ч.к.
41,5% 8,5% 8,5% 41,5%
Итак, Т. Морган не мог сказать, что расщепление идет по Г. Менделю (тогда бы доля каждого фенотипа равнялась 25%), но не наблюдалось и полного сцепления (тогда бы было всего два фенотипа, по 50% каждого). Числовые соотношения при этом скрещивании ближе к сцепленному наследованию. Поэтому такое наследование Т. Морган
назвал неполным сцеплением.
Причину неполного сцепления Т. Морган объяснил явлением кроссинговера, а особи, сочетающие признаки обоих родителей (серое тело, короткие крылья и чёрное тело, длинные крылья) назвал кроссоверными. Таких мух было 17%. Кроссинговер сопровождает образование любой гаметы, но при образовании разных гамет он происходит в разных участках данной пары хромосом. На участке, расположенном между генами (А) и (В), кроссинговер происходит при образовании 17% гамет, поэтому и кроссоверных особей 17%.
На основании своих работ Т. Морган сформулировал закон: "Сила сцепления генов обратнопропорциональна расстоянию между ними". Из закона следует, что величина кроссинговера зависит от силы сцепления генов: чем сила сцепления генов больше, тем меньше величина
кроссинговера. В настоящее время в генетике используется единица измерения расстояния между генами в хромосоме – морганида. Одна морганида равна 1% кроссоверных особей. Наличие в потомстве 50% и более кроссоверных особей говорит о независимом (менделевском) наследовании признака.
Примечание
Кроссоверные особи не появлялись в потомстве гетерозиготных самцов дрозофилы, т.к. при сперматогенезе у них не происходит кроссинговер.
3. Линейное расположение генов в хромосомах. Генетические и цитологические карты хромосом.
Изучение наследования сочетаний других признаков показало, что процент кроссоверного потомства для каждой пары признаков всегда один и тот же, но он различается для разных пар признаков. Это наблюдение стало основанием для заключения, что гены в хромосомах расположены в линейном порядке.
Генетические карты
Они строятся с учётом процента кроссоверных потомков. Чем ближе расположены два гена в хромосоме, тем меньше вероятность кроссинговера между ними, и, следовательно, меньше процента кроссоверных потомков. Больший процент рекомбинантных потомков в анализирующем скрещивании говорит о том, что гены расположены в хромосоме дальше друг от друга. Поэтому, определяя процент кроссоверных по различным признакам потомков, можно построить генетическую карту хромосомы.
За единицу расстояния и между генами принимается 1% кроссоверных особей, или 1 Сантиморганида.
Примечание
Если расстояние между генами больше 50 Сантиморганид, то говорят о независимом наследовании данного признака.
Генетические карты человека могут оказаться очень полезными в развитии медицины и здравоохранения. Уже в настоящее время знание о локализации гена на определённой хромосоме используется при диагностике ряда тяжёлых наследственных заболеваний человека. В будущем появится возможность для генной терапии, т. е. исправления структуры или функции гена.
Цитологические карты хромосом
Они показывают расположение генов в хромосоме как в цитологической структуре. При этом учитываются все гены, а не только гены, контролирующие признаки организма.
Цитологические карты составляются на основе дифференциальной окраски хромосом.
4. Механизм генетического определения и дифференцировки пола в развитии.
Диплоидный набор соматических клеток человека представлен 46 хромосомами или 23 парами хромосом. Из 46 хромосом 44 хромосомы (22 пары) одинаковы у мужского и женского организмов. Это аутосомы (обозначаются буквой А). Одна пара хромосом разная. Это половые хромосомы. У мужчин они представлены одной палочковидной хромосомой и одной хромосомой в виде крючка, напоминающего букву Y. Так эту хромосому и назвали, а вторую назвали Х-хромосомой. У женщин обе половые хромосомы палочковидные, т.е. Х-хромосомы. Тогда хромосомный набор соматических клеток можно представить так:
44 А + XX – клетки женского организма; 44 А + ХY – клетки мужского организма;
Хромосомный набор половых клеток выглядит так:
22 А + Х ; 22 А + Х – женские гаметы;
22 А + Х; 22 А +Y – мужские гаметы.
Следовательно, мужской пол продуцирует гаметы, отличающиеся по содержанию половых хромосом. Это гетерогаметный пол, женский пол – гомогаметный, т.к. продуцирует гаметы, содержащие одинаковые половые хромосомы.
Схема генетического определения пола человека (тип ХY). При этом типе наследования пола самец имеет одну Х-хромосому и одну Y- хромосому, а самка – две Х-хромосомы.
Р 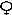 ХХ х
ХХ х 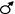 ХY Р 44 + ХХ х 44+ ХY
ХY Р 44 + ХХ х 44+ ХY

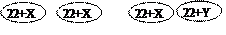 Г
Г
F1 ХХ, ХХ, ХY, ХY F1 44+ХХ, 44+ХХ, 44+ХY, 44+ХY
ж ж м м ж ж м м
Генетический пол ребенка определяется в момент оплодотворения и зависит от содержания в сперматозоиде хромосомы X или У. Таким образом, пол потомка определяет гетерогаметный родитель. Так как Х-сперматозоидов и Y-сперматозоидов образуется одинаковое количество, то теоретически соотношение полов = 1:1.
Кроме рассмотренного типа наследования пола, существуют другие и типы.
Тип ХО, или тип (кузнечик, клопы).
При этом типе наследования пола самец имеет одну Х-хромосому, а самка – две Х-хромосомы.

|
ХХ х ХО
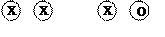 Г
Г
F1 ХХ , ХХ, ХО, ХО соотношение полов = 1:1, определяет пол потомков самец.
Тип Z W – самка гетерогаметна, а самец – гомогаметен (птицы, бабочки).
Р Z W х  ZZ
ZZ
 Г
Г
F1 ZZ, ZW, ZZ, ZW соотношение полов = 1:1, определяет пол потомка самка.
Тип ♀ 2 n, ♂ 1 n, половые хромосомы отсутствуют (пчёлы, муравьи)
Этот тип наследования пола связан с явлением партеногенеза.
Вывод: у всех организмов, кроме размножающихся партеногенезом, наследование пола идет по менделевским закономерностям.
Переопределение пола.
Пол будущего организма определяется в момент оплодотворения, но формирование конкретного фенотипа во многом зависит от внешней среды. В природе и в эксперименте возможно переопределение пола.
В морях у червя Bonellia личинки не дифференцированы по полу. Если личинки свободно плавают в воде, то превращаются в самок. Если личинки прикреплены к хоботку самки, то превращаются в самцов.
В тихом океане обитают рыбки Labroides, живущие стайками из самок и одного самца. Каждый из членов такой семьи, кроме самца, постоянно находится в состоянии стресса, источником которого является самец. При этом уровень напряженности различается от самки к самке, так что можно выделить α- самок, β- самок и γ-самок и т.д. Гибель самца вызывает превращение α-самки (главной самки, сбрасывающей напряжение) в самца с полноценными семенниками. Описанное переопределение зависит от уровня гормонов в организме, выделяемых клетками надпочечников.
У крокодилов в зависимости от температуры инкубации яиц могут появляться или самцы или самки. При температуре инкубации 28-290 С вылупляются самцы; при температуре инкубации 31-330 С вылупляются самки.
Из примеров, рассмотренных выше, следует, что генотип особи заключает в себе информацию о возможности формирования признаков того или иного пола, которая реализуется лишь при определенных условиях индивидуального развития. Изменение этих условий может стать причиной переопределения признаков пола. При этом у организмов разных видов относительное значение генетических и средовых факторов не одинаково: у одних видов определяющим фактором является среда, у других – наследственная программа.
5. Наследование признаков, контролируемых генами Х- и У-хромосом человека.
Х
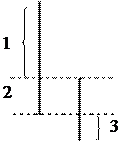 |
Y
В половых хромосомах имеются гомологичные и негомологичные участки:
1. участок Х-хромосомы, не имеющий гомолога в Y-хромосоме.
2. гомологичный участок Х и Y-хромосом;
3. участок Y-хромосомы, не имеющий гомолога в Х-хромосоме.
Наследование признаков, контролируемых генами Х- и У-хромосом человека называется наследованием, сцепленным с полом. В половых хромосомах находятся гены, которые отвечают за развитие как нормальных, так и патологических признаков.
Выделяют 3 типа наследования признаков сцепленных с полом.
1) тип Х. Признаки, наследующиеся по этому типу, контролируются генами, которые лежат в участке Х-хромосомы, не имеющий гомолога в Y-хромосоме (гемофилия, дальтонизм, отсутствие потовых желез).
2) тип У (голандрический). Признаки, наследующиеся по этому типу, контролируются генами, которые лежат в У-хромосоме. Признак передается строго от отца к сыну (некоторые заболевания почек, волосатость ушной раковины – гипертрихоз, развитие половых желёз по мужскому типу).
3) тип ХУ. Признаки, наследующиеся по этому типу, контролируются генами, которые лежат в гомологичных участках Х и Y-хромосом. Например, волчья пасть, заячья губа.
Пример: наследование гемофилии.
Это рецессивный признак, сцепленный с полом (с Х-хромосомой). Если женщина здорова, но в одной из Х-хромосом имеет ген гемофилии, она называется носительницей (ХН Хh). Она выходит замуж за здорового мужчину, но их дети (мальчики) могут быть больными.
Р Х H Xh x XH Y
 Г
Г
F1 XH XH , XH Xh , XH Y , Xh Y
Вероятность рождения в этом браке больных детей 25%, но в случае рождения мальчика вероятность повышается до 50%.
6. Хромосомная теория, основные положения.
Закономерности наследственности, открытые Г.Менделем, углубленные Т.Морганом и другими учёными, известны под общим названием: "хромосомная теория наследственности". Это учение о локализации генов в хромосомах, утверждающее, что преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Общие положения хромосомной теории наследственности:
1. гены находятся в хромосомах. Каждая пара хромосом есть группа сцепления генов. Число групп сцепления равно гаплоидному числу хромосом.
2. каждый ген в хромосоме занимает определённый участок. Гены в хромосомах расположены линейно.
3. между гомологичными хромосомами может происходить кроссинговер и обмен аллельными генами.
4. частота кроссинговера пропорциональна расстоянию между генами. Чем дальше гены расположены друг от друга, тем кроссинговер между ними происходит чаще.
Хромосомная теория наследственности была подтверждена при изучении генетических механизмов определения пола у животных и при изучении сцепленного наследования.
ЛЕКЦИЯ 7 Генотип и фенотип. Взаимодействие аллельных и
Дата добавления: 2016-07-18; просмотров: 3757;
Поиск по сайту
Узнать еще
- Алкоголь и его влияние на физическое и психическое здоровье человека. Профилактика
- Анатомо-физиологические аспекты потребностей человека. Задачи изучаемого предмета.
- Бактерии играют важную роль в жизни человека.
- Биомасса, или живое вещество биосферы. Закономерности распространения биомассы в биосфере, тенденции ее изменения под влиянием деятельности человека.
- В. Трансформация, вызванная генами-супрессорами
- Взаимодействия между генами в генотипе
- Виды ионизирующих излучений, их свойства и воздействие на организм человека.
- Виды человека. Расселение древнейших людей.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
