Нуклеиновые кислоты
Нуклеиновые кислоты впервые были выделены Ф.Мишером в 1869 г. из ядер клеток гноя, а сам термин предложен А.Косселем в 1889 г.
К нуклеиновым кислотам относят высокополимерные соединения, распадающиеся при гидролизе на пуриновые и пиримидиновые основания, пентозу и фосфорную кислоту. Нуклеиновые кислоты содержат С, Н, О большое количество Р (8-10%) и N (15-16%).
Значение нуклеиновых кислот для живых организмов заключается в обеспечении хранения, реализации и передачи наследственной информации.
| Нуклеотиды |
Нуклеотид — мономер нуклеиновых кислот. Молекула нуклеотида состоит из трех частей: азотистого основания, пятиуглеродного сахара (пентозы) и фосфорной кислоты (рис. 270).
| Состав нуклеотидов Азотистые основания |
Азотистые основания являются главной частью нуклеотида. Они имеют циклическую структуру, в состав которой наряду с атомами углерода входят атомы других элементов, в частности азота. За присутствие в этих соединениях атомов азота они и получили название азотистых, а поскольку они обладают щелочными свойствами — оснований.
Азотистые основания нуклеиновых кислот относятся к классам пиримидинов и пуринов.
Пиримидиновые основания являются производными гетероциклического соединения — пиримидина, имеющего в составе своей молекулы одно кольцо. К наиболее распространенным пиримидиновым основаниям относятся урацил, тимин, цитозин.
Пуриновые основания являются производными бициклического гетероцикла — пурина, имеющего два кольца: шестичленное и пятичленное.
К пуриновым основаниям относятся аденин и гуанин.
Во всех клетках — прокариотических и эукариотических — в состав нуклеиновых кислот входят эти пять основных азотистых оснований.
| Пятиуглеродный сахар |
Помимо азотистых оснований в образовании нуклеотидов принимают участие углеводный компонент, который представлен двумя сходными моносахаридами: рибозой или дезоксирибозой, относящихся к пентозам.
| Фосфорная кислота |
Третьим компонентом нуклеотидов является остаток фосфорной кислоты — фосфат. Именно наличие фосфата придает нуклеиновым кислотам свойства кислот.
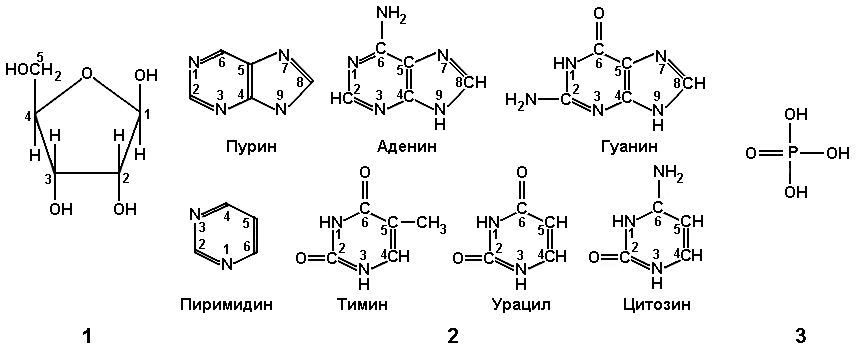 Рис. 270. Компоненты нуклеотидов:
1 — пятиуглеродный сахар; 2 — азотистые основания; 3 — фосфорная кислота.
Рис. 270. Компоненты нуклеотидов:
1 — пятиуглеродный сахар; 2 — азотистые основания; 3 — фосфорная кислота.
|
| Образование нуклеотидов |
Как отмечалось ранее, нуклеотиды являются мономерами нуклеиновых кислот. Биосинтез нуклеотидов является первым этапом биосинтеза нуклеиновых кислот. Они их непосредственные предшественники.
Образование нуклеотида происходит в два этапа. На первом этапе в результате реакции конденсации образуется нуклеозид — комплекс азотистого основания с сахаром. На втором этапе нуклеозид подвергается фосфорилированию. При этом между остатком сахара и фосфорной кислотой возникает фосфоэфирная связь. Таким образом, нуклеотид представляет собой нуклеозид, соединенный с остатком фосфорной кислоты (рис. 271).
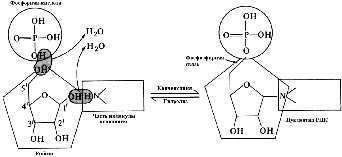 Рис. 271. Образование нуклеотида.
Рис. 271. Образование нуклеотида.
|
Названия нуклеотидов отличаются от названий соответствующих оснований. И те, и другие принято обозначать заглавными буквами:
Таблица 6.
Виды нуклеотидов:
| Азотистое основание | Нуклеотид | Обозначение |
| Аденин | Адениловый | А |
| Гуанин | Гуаниловый | Г или G |
| Цитозин | Цитидиловый | Ц или С |
| Тимин | Тимидиловый | Т |
| Урацил | Уридиловый | У или U |
| Функции нуклеотидов |
Нуклеотиды являются мономерами, из которых построены полимерные цепи нуклеиновых кислот, они входят в состав важных коферментов (НАД, НАДФ, ФАД, КоА).
| Образование ди- и полинуклеотидов |
Динуклеотид представляет собой соединение, состоящее из остатков двух нуклеотидов. При конденсации двух нуклеотидов между 3'-углеродом остатка сахара одного нуклеотида и остатком фосфорной кислоты другого возникает сложноэфирная связь. Таким образом, остатки сахаров двух
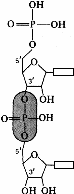 Рис.272. Образование динуклеотида
Рис.272. Образование динуклеотида
|
Возникновение фосфодиэфирных мостиков между 3' и 5'-углеродами остатков сахаров может происходить многократно. В результате образуются неразветвленные полинуклеотидные цепи. Один конец полинуклеотидной цепи заканчивается 5'-углеродом (его называют 5'-концом), другой –3'-углеродом (3'-концом).
| Полинуклеотиды |
В зависимости от углеводного компонента нуклеотидов, различают два класса нуклеиновых кислот:
© рибонуклеиновые кислоты (РНК), содержащие рибозу;
© дезоксирибонуклеиновые кислоты (ДНК), содержащие дезоксирибозу.
Нуклеотиды ДНК называют дезоксирибонуклеотидами, РНК — рибонуклеотидами.
| Дезоксирибонуклеиновая кислота |
Молекулы ДНК являются полимерами, мономерами которых являются дезоксирибонуклеотиды, образованные:
© остатком пятиуглеродного сахара — дезоксирибозы;
© остатком одного из азотистых оснований:
¨ пуриновых — аденина, гуанина;
¨ пиримидиновых — тимина, цитозина;
© остатком фосфорной кислоты.
| Структура молекулы ДНК |
ДНК представляет собой двойную спираль. Ее молекула образована двумя полинуклеотидными цепями, спирально закрученными друг около друга, и вместе вокруг воображаемой оси (рис. 273). Цепи ДНК антипараллельны (разнонаправлены), то есть против 3'-конца одной цепи находится 5'-конец другой. На периферию молекулы обращен сахаро-фосфатный остов, образованный чередующимися остатками дезоксирибозы и фосфатными группами. Внутрь молекулы обращены азотистые основания.
Диаметр двойной спирали ДНК — 2 нм, шаг общей спирали, на который приходится 10 пар нуклеотидов — 3,4 нм. Длина молекулы — до нескольких десятков и даже сотен микрометров. Молекулярный вес составляет десятки и сотни миллионов (для двойной спирали). В ядре клетки человека общая длина ДНК около 2м.
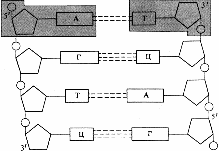 Рис. 273. Схематичное изображение развернутых цепей ДНК.
Рис. 273. Схематичное изображение развернутых цепей ДНК.
|
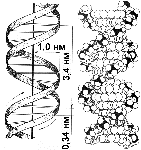 Рис.274. Трехмерная модель молекулы ДНК.
Рис.274. Трехмерная модель молекулы ДНК.
|
Трехмерная модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. американским биологом Дж.Уотсоном и английским физиком Ф.Криком (рис. 274). За свои исследования они были удостоены Нобелевской премии.
Полинуклеотидные цепи в молекуле ДНК удерживаются друг около друга благодаря возникновению водородных связей между азотистыми основаниями. Спаривание нуклеотидов не случайно, в его основе лежит принцип комплементарного взаимодействия пар оснований: против аденина одной цепи всегда располагается тимин на другой цепи, а против гуанина одной цепи — всегда цитозин другой, то есть аденин комплементарен тимину, а гуанин — цитозину (рис. 275). Комплементарностью называют способность нуклеотидов к избирательному соединению друг с другом.
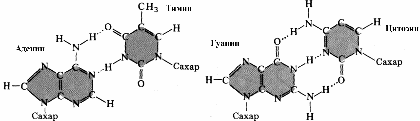 Рис. 275. Спаривание азотистых оснований.
Рис. 275. Спаривание азотистых оснований.
|
Комплементарность обеспечивается:
© взаимодополнением пространственных конфигураций молекул азотистых оснований;
© количеством водородных связей, возникающих между азотистыми основаниями (три водородные связи между гуанином и цитозином и две водородные связи между аденином и тимином).
Комплементарность пуриновых и пиримидиновых азотистых оснований обеспечивает одинаковое по всей длине двойной спирали расстояние между цепями.
Э.Чаргафф, обследовав огромное количество образцов тканей и органов различных организмов, выявил следующую закономерность: в любом фрагменте ДНК содержание остатков гуанина всегда точно соответствует содержанию цитозина, а аденина — тимину. Это положение получило название "правила Чаргаффа":
А + Г
А = Т; Г = Ц или ——— = 1
Ц + Т
Дж.Уотсон и Ф.Крик воспользовались этим правилом при построении модели молекулы ДНК.
Последовательность нуклеотидов одной цепи определяет последовательность нуклеотидов другой, поэтому две цепи молекулы ДНК комплементарны друг другу.
| Самоудвоение молекулы ДНК |
Одним из уникальных свойств молекулы ДНК является ее способность к самоудвоению — воспроизведению точных копий исходной молекулы. Благодаря этой способности молекулы ДНК, осуществляется передача наследственной информации от материнской клетки дочерним во время деления. Процесс самоудвоения молекулы ДНК называют репликацией.
Репликация — сложный процесс, идущий с участием ферментов (ДНК-полимераз) (рис. 276). Репликация осуществляется полуконсервативным способом, то есть под действием ферментов молекула ДНК раскручивается и около каждой цепи, выступающей в роли матрицы, по принципу комплементарности достраивается новая цепь. Таким образом, в каждой дочерней ДНК одна цепь является материнской, а вторая — вновь синтезированной. Раскручивание молекулы происходит на небольшом отрезке (несколько десятков нуклеотидов), называемом репликативной вилкой. После окончания синтеза дочерних цепей ДНК на данном участке и соединения их с материнскими раскручивается новый отрезок, и цикл репликации повторяется. Таким образом, репликативная вилка перемещается вдоль молекулы, пока не дойдет до точки окончания синтеза.
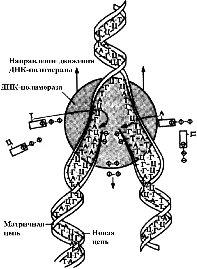 Рис. 276. Репликация ДНК.
Рис. 276. Репликация ДНК.
|
Во время репликации энергия молекул АТФ не расходуется, так как для синтеза дочерних цепей при репликации используются не дезоксирибонуклеотиды (содержат один остаток фосфорной кислоты), а дезоксирибонуклеозидтрифосфаты (содержат три остатка фосфорной кислоты). При включении дезоксирибонуклеозидтрифосфатов в полинуклеотидную цепь два концевых остатка отщепляются, и освободившаяся энергия используется на образование сложноэфирной связи между нуклеотидами.
Дата добавления: 2016-05-30; просмотров: 3749;
Поиск по сайту
Узнать еще
- А. ОДНООСНОВНЫЕ ПРЕДЕЛЬНЫЕ КИСЛОТЫ
- АМИДЫ СУЛЬФАНИЛОВОЙ КИСЛОТЫ (СУЛЬФАНИЛАМИДНЫЕ ПРЕПАРАТЫ)
- Б. ОДНООСНОВНЫЕ НЕПРЕДЕЛЬНЫЕ КИСЛОТЫ
- Биополимеры: белки, нуклеиновые кислоты.
- В норме аминокислоты, всосавшиеся в кровь из кишечника, циркулируют в крови 5 — 10 мин и очень быстро поглощаются печенью и частично другими органами (почками, сердцем, мышцами).
- В. ДВУХОСНОВНЫЕ КИСЛОТЫ ЖИРНОГО РЯДА
- Возбуждающие аминокислоты
- ВОПРОС №2. ПОЛУЧЕНИЕ МОЛОЧНОЙ КИСЛОТЫ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
