Рецепторы для цитокинов
Рц для цитокинов по структуре делят на 4 семейства:
1) семейство Рц для цитокинов класса I — семейство Рц для гемопоэтинов — гетеродимеры, гетеротримеры;
2) семейство Рц для цитокинов класса II — Рц для интерферонов и интерфероноподобных цитокинов — гомодимеры;
3) семейство Рц типа TNFR — мономеры, тримеризующиеся при связывании лигандами;
4) семейство Рц для хемокинов — 7–членная трансмембранная «гармошка».
Кроме того, ряд вирусных белков на поверхности клеток связывает некоторые цитокины. Например, белки цитомегаловируса связывают такие хемокины, как RANTES, MCP–1, MIP–1a и 1b. Молекулы гликозаминогликанов на поверхности клеток эндотелия связывают хемокины, например RANTES. Такое связывание не вызывает проведение сигнала внутрь эндотелиальных клеток. Его предназначение иное: RANTES на поверхности эндотелия связывает за Рц «для себя» лейкоциты из потока крови, что обеспечивает их экстравазацию и попадание в очаг воспаления в ткани.
Рц из семейства гематопоэтиновых представляют собой гетеродимерные молекулы. Это семейство включает: b и g–цепи Рц для ИЛ–2; Рц для ИЛ–3,4,5,6,7,9 и 15; Рц для GM–CSF; Рц для эритропоэтина; Рц для гормона роста. У Рц для ИЛ–3 и 5, GM–CSF одинаковая b–цепь. У Рц для ИЛ–2,4,7,9 и 15 одинаковая a–цепь. Есть варианты Рц для ИЛ–2. Первый — высокоаффинный (связывает лиганд с kd 10–11 М) состоит из 3 цепей — a (CD25, мол. масса 55 кДа), b (мол. масса 75 кДа, CD122) и g (мол. масса 64 кДа). Как гетеротример он экспрессирован на T–лимфоцитах памяти, некоторых B–лимфоцитах и NK. Второй вариант Рц — низкоаффинный Рц для ИЛ–2 — гетеродимер ИЛ–2Rbg, связывающий лиганд с kd 10–9 М.
Рц из семейства класса II — это Рц для интерферонов и ИЛ–10. Они являются гомодимерными трансмембранными молекулами.
Рц из семейства Рц для фактора некроза опухолей (TNFR) состоят из одной трансмембранной полипептидной цепи, имеющей два варианта по молекулярной массе — 55 или 75 кДа и включают TNFR–I и II, молекулу CD40, Fas (CD95), CD30, CD27, Рц для фактора роста нервов (NGF–R).
Рц семейства хемокиновых по структуре являются трансмембранной 7–слойной «гармошкой» и имеют множество конкретных вариантов.
Схема структуры названных семейств Рц к цитокинам показана на рис. 7.5.
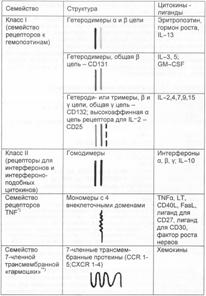
Рис. 7.5. Структурные семейства рецепторов для цитокинов. * — при функционально значимом связывании с лигандами Рц этого семейства тримеризуются в мембране; ** — к этому же структурному семейству относятся фоторецепторы — родопсин и бактериородопсин, а также Рц для анафилотоксинов комплемента (C5a, CЗa, C4a) и Рц для бактериальных пептидов с формилметионином (fMet) на N–конце.
Приведём более подробный список известных на сегодня Рц для хемокинов семейства CC, а также молекул, не являющихся только Рц для цитокинов, но способных связывать те или иные конкретные цитокины (табл. 7.9). Эта информация в определённых аспектах имеет большое не только научное, но и практическое значение. Например, один из вариантов Рц для хемокинов CCR5 оказался значимым кофактором для проникновения ВИЧ–1 в клетки. Вероятно, он не единственный.
Таблица 7.10. Рецепторы для хемокинов семейства CC
| Рецептор | Лиганды |
| CCR1 | MIP–1 |
| CCR2 | MCP–1, MCP–3, MCP–4 |
| CCR3 | Эотаксин, эотаксин–2/МРIF–2, RANTES, MCP–2, MCP–3, MCP–4, MIP–5, Lkn–1 |
| CCR4 | TARC |
| CCR5 | MIP1– |
| CCR6 | Exodus–1/LARC/MIP–3 |
| CCR7 | ELC/MIP–3 |
| CCR8 | I–309 |
| CCR10 | MCP–1, MCP–3, RANTES, MCP–4 |
| CMV US28 | MCP–1, RANTES, MIP–1 |
| DARC | RANTES и другие хемокины семейств CC и CXC |
| Гликозаминогликаны | RANTES и др. Это «несигнализирующий» Рц |
Примечание. CCR — Рц для хемокинов семейства CC; CMV US28 — белок цитомегаловируса; DARC — Аг группы крови Duffy, эту же молекулу использует малярийный плазмодий для проникновения в эритроциты, DARC связывает многие хемокины не только семейства CC, но и CXC; Lkn–1 — лейкотактин; MPC — моноцитарный хемоаттрактантный белок; MPIF — миелоидный ингибиторный фактор; TARC — thymus and activation regulated chemokine; LARC — liver and activation regulated chemokine; MIP — макрофагальный воспалительный белок.
При проведении сигналов от разных Рц для различных цитокинов, тем не менее, происходят общие внутриклеточные события. Цитоплазматические участки Рц для цитокинов ассоциированы с киназами определённого семейства, называемого Janus.
Название Janus происходит от имени двуликого древнеримского бога Януса и объясняется тем, что в молекулах киназ этого семейства обязательно присутствуют два домена: один является собственно ферментом — киназой (JH1, в активном центре молекулы содержатся АК–остатки KE/DYY); второй — псевдокиназный домен (JH2), осуществляющий регуляторные в отношении киназной активности функции. Всего в молекуле Janus 7 доменов, 5 из которых обеспечивают связывание с лигандами.
Известно 4 члена этого семейства, близких по структуре: Jak–1, Jak–2, Jak–3, Tyk–2. Их молекулярная масса составляет от 110 тыс. до 140 тыс. Эти киназы в клетке ассоциированы с разными цепями Рц для цитокинов. После связывания Рц с цитокином киназы Janus фосфорилируют внутриклеточные участки цепей Рц по остатку тирозина, после чего к этим участкам через SH2–домен могут присоединяться молекулы из семейства STAT (signal transducers and activators of transcription) — проводники сигналов и активаторы транскрипции. Затем те же киназы Janus фосфорилируют молекулы STAT по остатку тирозина. Фосфорилированные STAT отделяются от внутриклеточных цепей Рц для цитокина, димеризуются (гомо- или гетеро-) и в виде димера физически перемещаются в ядро, где вступают в связь с ДНК и активируют транскрипцию. В ДНК димеры STAT находят полупалиндромные последовательности нуклеотидов ТTN5–6АА и «садятся» на них «верхом». Для того чтобы началась транскрипция с ДНК, необходимо участие ещё так называемых коактиваторных белков, которыми могут быть, например, Рц для глюкокортикоидов, белки N–Myc, СВР/р300 и др. Показано, что онкопротеин аденовирусов Е1А способен блокировать функции STAT1 (пример механизма повреждения иммунных функций при аденовирусных инфекциях).
Описано 7 молекул STAT: 1, 2, 3, 4, 5а, 5b, 6. Их молекулярная масса составляет от 84 тыс. до 113 тыс. N–концевые домены молекул STAT участвуют в их димеризации. Существуют также спиральный домен, центральный ДНК–связывающий домен, SН2–домен и C–концевой домен (он обеспечивает собственно активацию транскрипции с ДНК). В молекулах STAT существуют консервативные остатки тирозина. Их фосфорилирование необходимо для димеризации. C–концевой домен фосфорилируется по остатку серина.
Дата добавления: 2016-07-18; просмотров: 2496;
Поиск по сайту
Узнать еще
- A ... метка (без метки) на шатуне (стрелка) для 26.20b Измерение внутреннего диаметра
- Andante cantabile con espressione В. Моцарт. Соната для ф-п. № 8, ч. II
- Andantino con moto А. Бородин. Для берегов отчизны дальней
- Audit Trail - Материалы для проведения аудиторской проверки
- I. Установка для исследования сдвига фаз колебаний силы тока и напряжения с помощью компьютера и осциллографа-приставки
- I.10. Тесты для контроля знаний
- III. Порошки для стирки и мытья
- III. ТИТАН – МАТЕРИАЛ ДЛЯ СОВРЕМЕННОЙ СТОМАТОЛОГИИ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
