Зрительное опознание жертвы и хищника
Большинство хищников встречается с множеством разных видов, которые служат им добычей и которых необходимо отличать от других организмов. При этом чаще всего используются три признака - величина, движение и форма.
Когда хищники должны выбирать между особями-жертвами, отличающимися только размерами тела, они обычно хватают самую крупную. Эта стратегия наиболее эффективна в отношении затрат энергии. Однако по мере увеличения размера обычно наступает предел, за которым стимул больше не воспринимается как добыча. Например, когда обыкновенной жабе (Bufo bufo) предъявляют как «добычу» объекты разной величины, она реагирует положительно на экземпляры, размеры которых находятся в определенных пределах, но активно избегает более крупные стимулы (рис. 13.6).
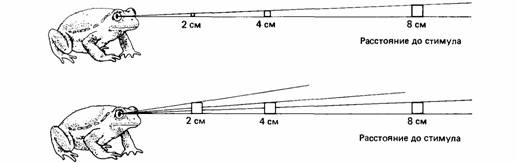
Как жаба судит о величине объекта? Проще всего делать это по величине изображения на сетчатке, измеряемого градусами угла зрения. Для объекта постоянной величины угол зрения меняется с расстоянием объекта от глаза. Близкие объекты выглядят крупнее, чем удаленные. Чтобы выбрать добычу определенной величины, жабе нужно оценить абсолютную величину видимого объекта, учитывая и его размеры на сетчатке, и расстояние до него. Жабы-повитухи (Alytes obstetricans] в процессе развития постепен-
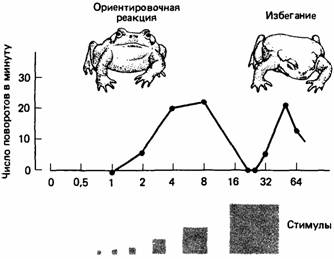
| Рис. 13.6. Жабы реагируют на маленькие движущиеся квадраты, ориентируясь в их направлении, но при определенных размерах квадрата эта реакция исчезает и большие квадраты вызывают реакцию избегания. (По Ewert, 1980.) |
но научаются выбирать добычу постоянного размера (Ewert, Burghagen, 1979). Сразу после метаморфоза они предпочитают искусственную приманку некоторого углового размера почти независимо от расстояния. Через шесть месяцев они ориентируются на объекты определенного абсолютного размера независимо от расстояния (рис. 13.7), т.е. за это время они как-то научаются включать суждения о расстоянии в свои оценки размеров. В некоторых случаях наблюдается склонность к более крупной добыче, но не из-за активного предпочтения, а потому, что она лучше различима. Так, радужные форели (Salmo gairdneri) чаще ловят крупных ракообразных, чем мелких, потому что крупные видны на большем расстоянии (Ware, 1972).
Для того чтобы узнать добычу, некоторым видам, например лягушкам и жабам, нужно, чтобы она двигалась. Обыкновенная каракатица (Sepia officinalis) в норме нападает только на движущихся креветок. Но если у каракатицы отнять только что пойманную и парализованную ею креветку, то это головоногое немедленно снова нападет на нее даже на неподвижную (Messenger, 1968). Некоторые хищники предпочитают добычу, которая движется беспорядочно. Так, личинки стрекоз предпочитают животных, совершающих зигзагообразные движения (Etienne, 1969), а солнечная рыба (Lepomis gibbosus) скорее нападет на искусственную рыбку, если та извивается, чем на плавно движущийся объект (Gandolfi et al., 1968). Иногда имеет значение соотношение формы и движения. Так, когда жабам предъявляют темную движущуюся полосу на белом фоне, они сразу же нападают на нее, когда полоса перемещается вдоль своей оси подобно червяку (рис. 13.8).
Узнавание добычи по форме - настолько сложное дело, что тут трудно сформулировать какие-нибудь общие правила. В ряде поведенческих работ Робинсон (Robinson, 1970) давал в неволе насекомых тамаринам (Saguinus geoffroyi) - восновном насекомоядным обезьянам Нового Света. В некоторых опытах им предъявляли богомолов - нормального, без головы, с двумя головами и т.д. Результаты показывают, что для узнавания добычи этим обезьянам важно было опознать голову. В опытах с палочниками в качестве приманки тамарины склонны были пренебрегать насекомыми без ног, но быстро хватали экземпляры с хорошо заметными ногами или маленькие палочки с прикрепленными к ним ножками насекомых. Таким образом, для опознания добычи тамаринам важны головы и конечности насекомых. Чтобы спастись от хищника, насекомые-жертвы выработали разнообразные защитные приспособления, скрывающие конечности и искажающие очертания тела. В опытах с абстрактными
|
| Рис. 13.7. Молодые жабы (вверху) оценивают размеры по углу зрения, а взрослые жабы (внизу) могут судить о размерах независимо от угла зрения. (По Ewert, 1980.) |
двумерными фигурами Робинсон обнаружил, что двусторонняя симметрия, вероятно, служит одним из признаков добычи, на который обычно ориентированы хищники.
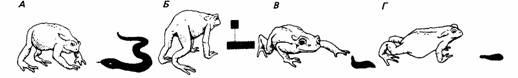
Те же принципы применимы и к узнаванию жертвами хищников. Так, например, силуэт ястреба, движимый над утятами или гусятами, вызывает реакцию страха, когда движется только в определенном направлении (рис. 13.9). Это объясняется тем, что короткая шея и длинный хвост характерны для ястреба, а длинная шея и короткий хвост - признаки летящего гуся (Tinbergen, 1951). Жабы избегают фигур, похожих на змею с поднятой головой (рис. 13.10). Пиявка, движущаяся толчками, воспринимается как добыча, если ее передняя присоска находится на почве, но если эта присоска поднята в воздух, жаба принимает пиявку за врага (Ewert, 1980).
Как мы видим, поведенческие исследования (см. обзор Ewert, 1980) показывают, что жабы хватают мелкие продолговатые объекты, расположенные горизонтально, но не реагируют на такие же объекты в вертикальном положении (рис. 13.8). Жаба обычно питается насекомыми, личинками, червяками и т. п. Ее хищническое поведение состоит из ориентации головы и туловища, зрительной фиксации добычи, захватывания ее в результате вытягивания шеи и языка, глотания и вытирания морды передними лапами (рис. 13.11). Чтобы вызвать хищническое поведение, нужен мелкий движущийся объект. На крупные движущиеся объекты жаба отвечает оборонительным поведением. Хотя для изучения сенсорных процессов могут быть применены разные чисто поведенческие методы, физиологическое исследование тоже может давать ценные сведения о деятельности органов чувств и о типе информации, которую они посылают в мозг. Но для того чтобы установить, как мозг использует такую информацию, тре-
| Рис. 13.8. Жабы реагируют положительно на прямоугольники, движущиеся вперед узкой стороной, и отрицательно на движущиеся вперед широкой стороной. (По Ewert, 1980.) |

| Рис. 13.9. Этот силуэт похож на ястреба, если его двигать в одном направлении, и на гуся, если его двигать в противоположном направлении. |
|
| Рис. 13.10. Реакции жабы на простые модели: А - змея; Б - абстрактный рисунок; В - пиявка с поднятой головой; Г - пиявка с опущенной головой. Первые три вызывают реакцию избегания, последняя исследуется. (По Ewert, 1980.) |
буется сочетание поведенческого и физиологического подходов. Такой подход был применен Эвертом и его коллегами в обширных исследованиях по опознанию добычи и врагов жабами.
Физиологические работы показывают, что некоторое опознание добычи происходит на уровне сетчатки. Леттвин и др. (Lettvin et al., 1959) отводили электрическую активность от зрительного нерва лягушки, когда объекты двигались в поле зрения. Они обнаружили четыре типа ответов, которые, по-видимому, соответствуют четырем типам ганглиозных клеток в сетчатке. Было установлено, что эти клетки являются детекторами: 1) неподвижной границы, 2) темного выпуклого движущегося объекта, 3) изменения контраста или движения и 4) затемнения. Жаба, очевидно, обладает тремя типами ганглиозных клеток (соответствующих типам 2, 3 и 4 у лягушки), аксоны которых в составе зрительного нерва идут в зрительную покрышку головного мозга (Ewert, Hock, 1972). Информация, поступающая в мозг, включает угловой размер и скорость движения объекта, степень контраста с фоном и общий уровень освещенности. Однако, чтобы распознать добычу, жабе этой информации недостаточно.
Методом дегенерации можно проследить путь волокон зрительного нерва в разные отделы головного мозга, включая зрительную покрышку и претектальную область таламуса (рис. 13.12). Сетчатка одного глаза проецируется топографически на поверхностные слои противоположной зрительной покрышки. Эта проекция имеет вид карты, на которой каждая область поля зрения соответствует определенной области зрительной покрышки. Электрическая стимуляция покрышки у свободно движущейся жабы приводит к ориентировочной реакции по направлению к соответствующей части поля зрения, т. е. вызывает такое поведение, как будто соответствующая часть зрительной покрышки стимулируется видом добычи. Электрическая стимуляция проекции сетчатки на таламической претектальной области вызывает у жабы реакцию избегания.
|
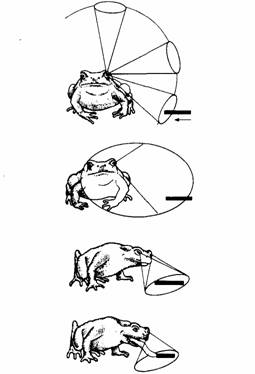
| Рис. 13.11. Обнаружение и схватывание добычи жабой. Сверху вниз: добыча попадает в боковое поле зрения животного; жаба поворачивается к добыче, подводя ближе к ней центр поля зрения, а в результате используя бинокулярное поле; затем жаба схватывает добычу языком. (По Ewert, 1980.) |
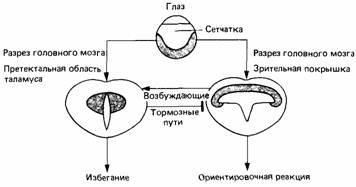
| Рис. 13. 12. Соотношение между частями головного мозга жабы, участвующими в регуляции избегания и в ориентировочной реакции на зрительно обнаруживаемые обьекты. (По Ewert, 1980.) |
Хирургическое разрушение части головного мозга (претектальной области таламуса) приводит к тому, что животные начинают хватать любой движущийся предмет. Разрушение зрительной покрышки уничтожает всякую реакцию на движущиеся стимулы, в том числе и поведение избегания. Эти данные побудили Эверта (Ewert, 1980) постулировать, что проекция сетчатки на претектальную область таламуса вызывает поведение избегания, но что для этого также требуется некоторый приток возбуждения от зрительной покрышки. Проекция сетчатки на зрительную покрышку служит основой реакции схватывания всех движущихся объектов, но реакция на крупные или похожие на врага объекты тормозится претектальной областью таламуса и схватываются только мелкие объекты.
Гипотеза Эверта подтверждается физиологическими работами по изучению электрической активности нейронов в зрительной покрышке и претектальной области таламуса в ответ на стимуляцию сетчатки и других связанных с этой функцией частей головного мозга (Ewert, 1980). Это исследование представляет большой интерес не только как демонстрация соответствующего участия сетчатки и мозга в фильтрации стимула, но так же как прекрасный пример того, чего можно достичь разумным сочетанием поведенческих и физиологических методов.
ДЛЯ ЗАПОМИНАНИЯ
1. Животные, обитающие при слабом освещении, часто обладают специальными зрительными приспособлениями - глазами цилиндрической формы, тапетумом и зрительными пигментами, - предназначенными для повышения чувствительности в этих условиях.
2. У животных, которые живут или активны в темноте, не позволяющей видеть окружающие предметы, глаза могут дегенерировать и зрение заменяют другие сенсорные системы. К ним относятся эхолокационная система летучих мышей, система слуховой локализации у сов и электрическая чувствительность у некоторых рыб.
3. Зрительное опознание добычи и хищников часто включает в себя обнаружение ключевых раздражителей, что позволяет животному быстро принимать решение и быстро реагировать.
Рекомендуемая литература
Gould J. L. (1980) The case for magnetic-field sensitivity in birds and
bees (such as it is). American Scientist, 68,256-267.
Lythgoe J.N. (1979) The Ecology of Vision, Clarendon, Oxford.
Дата добавления: 2016-06-18; просмотров: 2982;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
