Решите задачи (3 б.)
1. Синтез какого количества АТФ обеспечивает один цикл β-окисления жирной кислоты. По какой формуле можно рассчитывать количество молекул АТФ, образующихся при полном распаде до конечных продуктов насыщенной жирной кислоты. Сделайте расчет для лауриновой кислоты.
2. Сколько молекул макроэргов (АТФ) затрачивается на синтез одной молекулы нейтрального жира из глицерина и трех молекул жирных кислот? Ответ объясните.
3. Сколько молекул макроэргов (АТФ, ЦТФ) затрачивается на синтез одной молекулы лецитина из глицерина, двух молекул жирных кислот и холина. Ответ поясните.
Оцените знания, сверив вариант своего ответа с эталоном правильных ответов. Если Вы набрали от 34 – 17 баллов, значит Вы успешно изучили материал. Если Вы набрали менее 17 баллов, рекомендуем Вам ещё раз внимательно поработать над данной темой и ещё раз проверить себя, поработав с предлагаемыми заданиями.
Ответы к системе заданий по теме «Метаболизм липидов»
Часть А
1) г;
2) г;
3) д;
4) г;
5) д;
6) в;
7) б;
8) г;
9) д;
10) в;
11) б;
12) г;
13) д;
14) б;
15) б.
Часть Б
1.Активирование пальмитиновой кислоты:
Н4 Р2 О7
 + АТФ +НS~КоА АМФ
+ АТФ +НS~КоА АМФ
С15-Н31-СООН − − − − → С15Н31-СO~О-АМФ – – – – – – →
Пальмитат Ацил-КоА- Ациладенилат Ацил-КоА-синтетаза
Синтетаза
– – – –→ С15Н31-СO~S-КоА
Пальмитоил-КоА
(активированная пальмитиновая кислота)
Фермент: Ацил-КоА-синтетаза относится к классу лигаз (синтетаз).
2. β-окисление активированной пальмитиновой кислоты:
СН3
СН3 СН3 |
 | +НОН (СН2)12
| +НОН (СН2)12
( СН2)12 2Н (2 АТФ) (СН2)12 − − − − − − → |
| – – – – – → Еноил-КоА β НО - СН − − − − − − →
β СН2 Ацил-КоА-дегид- гидратаза |
| рогеназа (1) β СН (2) α СН2
α СН2 ФАД || |
| α СН СО~S−КоА
СО~S−КоА |
СО~S−КоА
Пальмитил-КоА Еноил-КоА β-Гидроксиацил-КоА
(ацил-КоА)
 2Н (3 АТФ)
2Н (3 АТФ)
СН3 СН3
− − − − → +HS~КоА |
β-Гидроксиацил-КоА- (СН2)12 − − − − − − → (СН2)10 + CH3CO~SKoA
дегидрогеназа (3) | Ацетил-КоА-ацил- |
НАД+ С=О трансфераза (4) СН2 Ацетил-КоА
| |
СН2 СН2
| |
СО−S−КоА СО~S−КоА
β-Кетоацил-КоА Миристил-КоА
(ацил-КоА)
Миристил-КоА вновь подвергается многократному (6 раз) β-окислению, ацетил-КоА вступает в цикл трикарбоновых кислот или используется для синтеза новых жирных кислот, холестерина, кетоновых тел, водород же поступает в цепи транспорта электронов. Ферменты 1 и 3 относятся к классу оксидоредуктаз, фермент 2 - к классу лиаз, фермент 4 - к классу трансфераз.
3.β-восстановление:
НАДФ+ СН3
 СН3 | Н2О
СН3 | Н2О
| +НАДФН(Н+) (СН2)12
(СН2)12 − − − − − − − − − − − − − − − − → | − − − − − − − − − −→
| 3-Оксоацил-АПБ- β СН-ОН 3-Гидроксиацил-АПБ-
С=О редуктаза (1) | дегидратаза (2)
| α СН2
СН2
О О
// //
C~S−АПБ C~S−АПБ
β-Оксоацил-АПБ
(β-кетопальмитил-АПБ) β-Гидроксиацил-АПБ
НАДФ+
 СН3 СН3
СН3 СН3
| +НАДФН (Н+) |
(СН2)12 (СН2)12
− − − −→ | − − − − − − − − − − − − → |
β СН Еноил-АПБ-редуктаза (3) СН2
|| |
α СН СН2
О
// О
С~S−АПБ //
Еноил-АПБ С~S−АПБ
Ацил-АПБ (пальмитил-АПБ)
Ферменты 1 и 3 относятся к классу оксидоредуктаз, а фермент 2- к классу лиаз.
4. Образование глицерина из белков:
Белки − − − → Аминокислоты − − − → Пируват − − − − →
Распад (кроме лейцина и лизина) Распад Реакция глюконеогенеза
+НАДН (Н+) НАД+
− − − − → Дигидроксиацетон − − − − − − − − → α - Глицерофосфат
фосфат Глицерол-3-фосфат-
дегидрогеназа
Образование глицерина из углеводов:
СН2-ОН +НАДН(Н+) НАД+
 | СН2-ОН
| СН2-ОН
С=О |
Глюкоза − − − − −→ | − − − − − − → НО−C−H
 Дихотомический СН2−О− Р Глицерол-3-фосфат |
Дихотомический СН2−О− Р Глицерол-3-фосфат |
распад дегидрогеназа СН2-О- Р
Дигидроксиацетон-
фосфат α -Глицерофосфат
(активный глицерин)
5.Синтез фосфатидилхолина:
СН2-О-СО-С13Н27 Н3РО4 СН2-О-СО-С13Н27
 | +Н-ОН |
| +Н-ОН |
С17Н23-СО-О-СН − − − −−−−−→ С17Н33-СО-О-СН
| Фосфатидатфосфатаза (1) |
СН2-О-Р СН2-ОН
Фосфатидная кислота Диацилглицерин
 + СН3
+ СН3
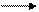 +ЦДФ-О-СН2-СН2-N СН3 ЦМФ
+ЦДФ-О-СН2-СН2-N СН3 ЦМФ
СН3 СН2-О-СО-С13Н27
|
− − − − − − − − − − − − − − − − → С17Н33-СО-О-СН
 Холинфосфат- | + СН3
Холинфосфат- | + СН3
рансфераза ( 2) СН2-О- Р - О-СН2-СН2-N - СН3 СН3
Фосфатидилхолин (лецитин)
Содержит миристиновую и олеиновую кислоты
Фермент 1 относится к классу гидролаз, а фермент 2 к классу трансфераз.
Часть С
1. Один цикл β-окисления жирной кислоты обеспечивает синтез 5 молекул АТФ, плюс 12 молекул АТФ возникает при распаде отщепляющегося ацетил-КоА в цикле трикаброновых кислот. Общее количество молекул АТФ, образующихся при полном распаде жирной кислоты с четным числом С-атомов, можно рассчитать по форме (n/2·17 – 6), где n –число углеродных атомов в жирной кислоте. Для лауриновой кислоты С11Н23СООН это количество составляет 96 молекул АТФ.
2. На синтез одной нейтрального жира из свободных глицерина и жирных кислот расходуется 4 молекулы АТФ: по 1 молекуле АТФ на активирование глицерина и каждой жирной кислоты.
3. На синтез одной молекулы лецитина (фосфатидилхолина) из свободных глицерина, двух жирных кислот и холина расходуется 5 микроэргических молекул (4 АТФ и ЦТФ): по 1молекуле АТФ на активирование глицерина и двух жирных кислот и по 1 молекуле АТФ и ЦТФ на активирование холина.
Заключение
Уже при кратком знакомстве с молекулярными основами жизни мы сталкиваемся с липидами. Объединение липидных веществ, принадлежащих к различным классам химических соединений, в одно большое семейство было сделано на основании их крайне важной роли в регуляторных процессах в живых организмах. Перечень выполняемых ими биологических функций огромен, и он увеличивается по мере изучения липидов. Поэтому для понимания сути многих физиологически важных процессов нужно иметь представления о липидах на таком же уровне, как о белках, нуклеиновых кислотах и углеводах.
Необходимость глубокого и детального изучения данного класса биологических соединений, механизмов процессов их обмена и регуляции связана с тем, что липиды широко используются в разных отраслях народного хозяйста, в промышленности, в медицине. Кроме того, для лечения целого ряда заболеваний современности (атеросклероз, ожирение и др.) и предотвращения их негативного влияния на функционирование организма необходимо понимание механизма их развития, что невозможно без знания нормальных процессов метаболизма липидов.
Учитывая все вышесказанное, в данном пособии подробно рассмотрены вопросы статической и динамической биохимии липидов, механизмов их регуляции и интеграции, а также основных нарушений их метаболизма. Излагаемый материал отражает основные достижения в области биохимии, опубликованные в учебной и научно-популярной литературе последних лет как отечественных, так и зарубежных авторов.
Обобщая материал, рассмотренный в пособии, можно сделать следующие выводы:
1. Липиды являются важнейшими биомолекулами, играющими исключительно важную роль в процессе функционирования живых организмов. Помимо каноническихфункций липидов (энергетическая, запасная, поставщик метаболитов и структурная), в последнее время выделены неканонические функции,связанные сучастием липидов в деятельности мембранного аппарата клетки (регуляция деятельности ряда гормонов и активности ферментов, влияние на процессы транспорта метаболитов и макромолекул, контроль реакций биологического окисления и энергетического обмена, связь с репликацией ДНК и ее матричной активностью, компартментализация обменных процессов в клетке вплоть до формирования мембранных структур, участие в межклеточных взаимодействиях и др.).
Специфические биологические функции разных групп липидов связаны с особенностями их строения; например, жиры являются главным источником энергии в организмы, в то время как полярные фосфолипиды являются важнейшими компонентами клеточных мембран.
2. Разнообразие и уровень липидов в клетках, тканях и органах определяются процессами липидного метаболизма (ЛМ), включающими их транспорт, поглощение, использование клетками, синтез, разрушение и выведение. Процессы липидного метаболизма происходят при участии множества белков с различными функциями, которые, как и кодирующие их гены, также являются компонентами системы липидного метаболизма. Метаболизм липидов - совокупность процессов их катаболизма и анаболизма, протекающих в клетках, тканях и субклеточных структурах организма. Одной из главных задач обмена липидов является обеспечение организма энергией. Реакции β-окисления и последующего окисления ацетил-КоА в цикле трикарбоновых кислот служат одним из основных источников энергии для синтеза АТФ по механизму окислительного фосфорилирования. Биосинтез жирных кислот и триацилглицеролов является необходимым для пополнения энергетических ресурсов организма.
Синтез и разрушение липидов происходят практически во всех тканях организма. Вместе с тем, ряд тканей выполняют специализированные функции. Так, поглощение экзогенных липидов происходит в стенках тонкого кишечника; запасание - в жировой ткани; выведение продуктов распада липидов - в кишечнике, почках, легких. Центральное место в обмене липидов занимает печень, в которой происходит пересечение путей метаболизма липидов, углеводов и белков. Здесь же синтезируется основная масса белков транспорта липидов, также продукты деградации липидов, выводящиеся из организма.
3. Интенсивность и направленность различных превращений липидов должны соответствовать потребности организма в энергетическом и пластическом материале. Поэтому крайне важными становятся вопросы регуляции обмена липидов на уровне организма с помощью нервной и эндокринной систем. При нарушении механизмов регуляции развиваются различные патологические отклонения, ведущие к развитию целого ряда заболеваний. Обмен липидов тесно связан с различных групп обменом соединений других классов, а метаболические пути метаболизма липидов являются частью общей метаболической сети, функционирующей в организме.
В последнее время биохимия липидов стремительно развивается. В частности, в последних исследованиях оказалось, что блокирование работы одного из сигнальных липидов может приводить к серьезным нарушениям в развитии нервной системы, начиная от торможения роста новых нервных окончаний и заканчивая гибелью нейронов.
В настоящее время наметилось также новое направление и в изучении обмена липидов. Оно касается достаточно энергично протекающего процесса межмембранного переноса липидов, особенно фосфолипидов, из митохондрий в эндоплазматическую сеть и обратно, из мембранной фракции клетки в липосомы, от липосом одного состава к липосомам другого состава, от внутреннего липидного слоя мембраны к внешнему и наоборот и т.п. Значение этого динамично протекающего обновления и видоизменения липидного состава мембран огромно, так как при его посредстве регулируется метаболическая активность мембранного аппарата клетки и субклеточных структур.
Дата добавления: 2016-06-09; просмотров: 3345;
Поиск по сайту
Узнать еще
- II. Основные задачи ГО
- II. Основные задачи службы торговли и питания
- NP-СЛОЖНЫЕ И ТРУДНОРЕШАЕМЫЕ ЗАДАЧИ
- NP-сложные и труднорешаемые задачи
- X. ситуационные задачи по 3 модулю
- Автоматизация технологического проектирования. Основные задачи и модели автоматизации технологического проектирования
- Автомобильной службы части (соединения) после получения задачи
- Алгоритм и этапы решения задачи

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
