Участие в программировании движений премоторных полей коры.
Поражение премоторных зон у человека не сопровождается параличом контралатеральных конечностей. Вместе с тем их основным симптомом является отчетливое нарушение двигательных навыков. Клинически это проявляется в том, что у больного изменяется почерк, машинистка теряет быстроту и плавность своей работы, квалифицированный рабочий не способен автоматизированно выполнять серию операций, входящих в привычный двигательный акт. В клинике такие симптомы описывают под названием «инертность двигательных стереотипов». Характерными являются невозможность больного переключаться, например, с сильных ударов на слабые при отстукивании ритма ладонью, а также своеобразное «заклинивание» движения, двигательная персеверация (рис. 5.23). Наиболее отчетливо нарушения проявляются в контралатеральной очагу руке, но при поражении левого (доминантного) полушария они часто проявляются в работе обеих рук. Клиницисты отмечают, что у таких больных сохранены как намерения выполнить движение, так и общий план его выполнения, однако в своей исполнительной части движение высвобождается из-под влияния двигательной программы.

. В тех случаях, когда поражение располагается в нижних отделах премоторной зоны левого (доминантного по речи у правшей),полушария, происходит нарушение речи. Нарушения проявляются при переходе больного от одной артикуляции к другой (при произнесении любого слова или словосочетания). Нарушение кинетической организации артикуляции является основой той формы моторной афазии, которую описал П. Брока (1861). Следует отметить, что нарушения у больных с поражениями нижних отделов левой премоторной зоны проявляются не только в устной речи, но и в письме. Нарушения письменной речи считают следствием нарушения плавности перехода от одного компонента слова к другому и патологической персеверации однажды написанного слова. Некоторые дополнительные сведения Вы найдете в главе 7 «Высшие функции нервной системы».
Регистрация активности отдельных нейронов показала, что нейроны премоторной коры низших обезьян активируются до выполнения движения, что может указывать на их связь с программированием этого движения. При этом в структуре нейронной реакции находит отражение направление, в котором будет производиться данное движение (в ответ, например, на условный сигнал). Изучение корреляции нейронных характеристик с параметрами движений рук у обезьяны приводит исследователей к мысли, что в премоторных нейронных сетях содержится своеобразная «библиотека» отдельных целенаправленных движений. Например, если обезьяна подносит рукой ко рту кусочек пищи, то рисунок нейронного разряда не зависит от траектории этого движения (например, из разных точек экстраперсонального пространства). Предполагают, что премоторная кора у приматов участвует в программировании целенаправленных движений (рук) в экстраперсональном пространстве (в пределах вытянутой руки).
Участие в двигательном контроле теменной коры. По современным данным, поле 5 теменной коры также принимает участие как в сенсорных, так и в моторных процессах. Поле 5 часто рассматривают как верхний этаж обработки кинестетической информации. Показано, что при локальном обратимом выключении (например, охлаждением) поля 5 у обезьян нарушались условные двигательные рефлексы, выполнение которых существенно зависело от сохранности проприоцептивного контроля, а при выключении поля 7 страдала также и зрительная обратная связь. У кошек глубокий распад условных двигательных рефлексов на зрительные сигналы происходил только при совместном разрушении полей 5 и 7, тогда как при изолированном разрушении каждого из теменных полей условные двигательные рефлексы заметно не страдали.
Данные, полученные при исследовании нейронных сетей, указывают, что нейроны моторной коры кодируют движение в координатах динамики движения и имеют дирекциональную), активность определяется направлением движения конечности) настройку. В отличие от этого нейронные сети поля 5 кодируют пространственную траекторию движения, и эти нейроны лишь незначительно модулируют свою активность при воздействии нагрузок на движущуюся руку. Отсюда был сделан вывод, что, нейронные сети моторной коры кодируют движение в координатах его динамики, тогда как нейроны теменной коры кодируют пространственную траекторию движения.
Кортико-руброспинальная система двигательного контроля.От красного ядра, которое залегает в толще среднего мозга, берет начало руброспинальный путь. Моторная область коры, откуда берет начало пирамидный тракт, проецируется ипсилатерально на задние 2/3 красного ядра. Сенсомоторная кора (у кошки задняя часть сигмовидной извилины) проецируется ипсилатерально на переднюю область мелкоклеточной части красного ядра.
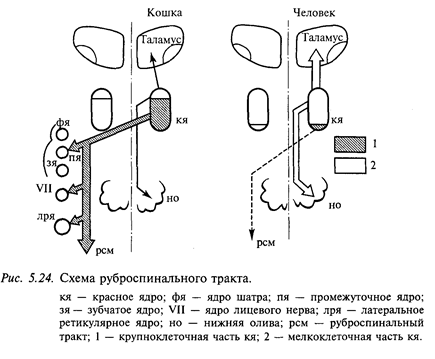
В ряду позвоночных животных структура красного ядра претерпевает существенные изменения (рис. 5.24). У рептилий и птиц оно однородно и содержит только крупные нейроны. У млекопитающих в дополнение к крупноклеточной (диаметр сомы 50-90 мкм) каудальной части ядра появляется ростральная часть, которая состоит преимущественно из мелких и средних нейронов (диаметр сомы 20-40 мкм). Соотношение этих двух разделов ядра варьирует, но у приматов увеличивается мелкоклеточная часть. Главный эфферентный путь красного ядра - руброспинальный тракт. После перекреста на уровне среднего мозга он спускается в мозговом стволе. В спинном мозге руброспинальный тракт идет в составе латеральных столбов (ростральнее латерального пирамидного тракта) и прослеживается до крестцовых сегментов спинного мозга. Основная часть афферентных проекций на мелкоклеточную часть красного ядра берет начало в зубчатом ядре мозжечка, а на мелкоклеточную часть - в промежуточных ядрах (круглом и пробковидном) мозжечка. Мелкоклеточное подразделение красного ядра образует восходящие проекции на вентролатеральное ядро таламуса, а нисходящие - на нижнюю оливу в продолговатом мозге. Увеличение размеров мелкоклеточной части и уменьшение крупноклеточной части красного ядра происходит на поздних стадиях эволюции и совпадает с появлением неомозжечковых структур и основной оливы. Регресс крупноклеточной части наиболее сильно выражен у человека и совпадает с неразвитостью руброспинального тракта.
Рассечение ствола мозга у четвероногих (кошка, собака) на уровне ростральное переднего двухолмия не вызывает больших изменений в мышечном тонусе. Однако если разрез проходит на интраколликулярном уровне (изолируется красное ядро), это приводит у четвероногих животных к ригидности (так называемая децеребрационная ригидность). Она развивается по гамма-типу, т. е. обусловлена повышенной импульсацией в гамма-мотонейронах. В отличие от ригидности по альфатипу эта ригидность пропадает после перерезки дорсальных корешков спинного мозга. Был сделан вывод, что крупноклеточная часть красного ядра осуществляет тормозное влияние на экстензоры постуральных мышц.
Опыты с электрической стимуляцией красного ядра показали, что преимущественно возбуждаются флексоры ипсилатеральной конечности. При стимуляции отдельных точек красного ядра прослеживается соматотопия.
В настоящее время есть основание предполагать, что обе системы двигательного контроля -кортико- и руброспинальные - организованы на уровне нейронного аппарата сегментарных механизмов спинного мозга в значительной степени сходно. Волокна обеих систем моносинаптически возбуждают в основном специализированные промежуточные нейроны латеральных участков серого вещества основания дорсального рога и нейроны промежуточного ядра и уже через эти переключения возбуждают мотонейроны.
Нейрофизиология ретикулоспинальных путей. Начало ретикулоспинальным путям дают нейроны на всем протяжении ретикулярной формации мозгового ствола. Эти пути спускаются в составе латеральных канатиков спинного мозга. Часть волокон в стволе мозга перекрещивается. Волокна ретикулоспинальной системы имеют различный диаметр и обеспечивают проведение возбуждения в широком диапазоне скоростей.
Облегчающие ретикулоспиналъные влияния. Американские нейрофизиологи Р. Райнс и Г. Мэгун (1946) установили, что слабое электрическое раздражение ретикулярной формации мозгового ствола и более ростральных отделов мозга усиливает двигательные реакции на раздражение моторной области коры и заметно облегчает коленный рефлекс (рис. 5.25). Эти эффекты опосредованы ретикулоспинальными волокнами, преимущественно идущими в составе латерального канатика. Область, из которой вызываются двусторонние облегчающие влияния, включает значительную часть ретикулярной формации продолговатого мозга, покрышку моста и среднего мозга, центральное серое вещество, субталамус и гипоталамус, а также срединное и интраламинарные ядра таламуса.
Облегчающая ретикулоспинальная система играет важную роль в развитии децеребрационной ригидности и спастичности. На децеребрированных кошках (перерезка мозгового ствола на межколликулярном уровне) было показано, что стимуляция участков облегчающей системы ниже уровня перерезки усиливает и без того увеличенные в таких условиях проприоцептивные рефлексы. Этот эффект развивается билатерально. Влияние облегчающей системы мозгового ствола на экстензорный тонус было убедительно показано на кошках с хронической спастичностью, которая развивалась в результате удаления двигательной области коры больших полушарий. Спастическое состояние, устраняющееся во время эксперимента на фоне наркоза, не только восстанавливалось, но существенно усиливалось при стимуляции облегчающей системы мозгового ствола вплоть до развития судорог.
Дата добавления: 2016-06-05; просмотров: 1793;
Поиск по сайту
Узнать еще
- II. Участие Республики Казахстан в международных договорах
- III. Исследование движений
- Амплитуда движений в ПДС у здорового человека
- Анализ, ошибки и оценки выполнения техники движений.
- Афферентный механизм произвольных движений afferentis — «приносящий»
- БИОЛОГИЧЕСКАЯ ЗАЩИТА ОТ ВОЗДЕЙСТВИЯ ЭЛЕКТРИЧЕСКИХ И МАГНИТНЫХ ПОЛЕЙ
- Биофизические принципы исследования электрических полей тканей и органов.
- Вертикальные движения воздуха в атмосфере. Причины и виды вертикальных движений.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
