НЕЙРОФИЗИОЛОГИЯ ЗРИТЕЛЬНОЙ СИСТЕМЫ
Зрение в жизни человека имеет огромное значение. Это основной сенсорный канал, который связывает его с внешним миром. Зрительная система человека устроена очень сложно. Благодаря зрению мы воспринимаем окружающий нас мир в объеме и красках, мы читаем и смотрим кино, телевизор. И это все зрение.
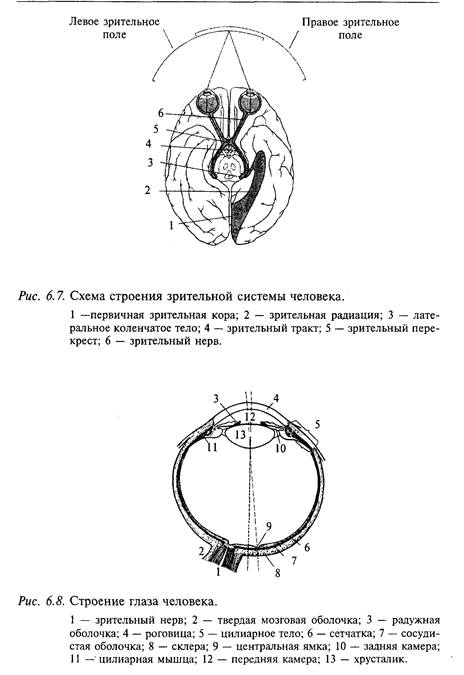
В зрительной системе человека можно выделить следующие уровни обработки сигналов. На периферии находится сетчатка. В ходе развития нервной системы сетчатка закладывается на самых ранних этапах развития (так называемые «глазные пузыри»). Поэтому есть все основания считать сетчатку «частью мозга, вынесенного на периферию». Следующий уровень обработки зрительной информации находится в таламусе - это наружное коленчатое тело. Аксоны нейронов наружного коленчатого тела проецируются в кору затылочного полюса больших полушарий (поля 17, 18, 19). Высший этап обработки зрительных сигналов происходит в ассоциативных полях коры больших полушарий. Схема зрительной системы приведена на рис. 6.7.
Строение глаза. Глаз человека имеет шарообразную форму (рис. 6.8). Вращение глазного яблока в глазнице осуществляется тремя парами мышц, которые иннервируются глазодвигательными черепными нервами (подробнее см. выше). Плотная наружная оболочка глаза образована непрозрачной склерой, которая на переднем полюсе переходит в прозрачную роговицу. Внутри глазного бокала находится сосудистая оболочка, содержащая кровеносные сосуды. Впереди сосудистая оболочка переходит в ресничное тело и далее в радужку. В радужке находятся гладкие мышечные волокна, степень напряжения которых определяет диаметр зрачка. При сокращении или расслаблении гладкой мускулатуры ресничного тела изменяется напряжение цинновых связок, от которых зависят радиус кривизны хрусталика и его преломляющая сила, т. е. аккомодация глаза. Пространство между хрусталиком и роговицей, называемое передней камерой, заполнено прозрачной жидкостью, между хрусталиком и сетчаткой - студенистой жидкостью, или стекловидным телом. Дно глазного бокала выстлано сетчаткой.
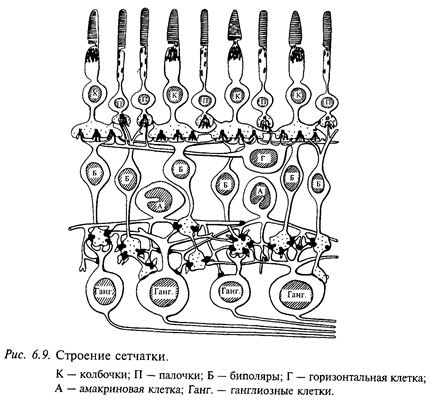
Сетчатка по своему строению и происхождению представляет собой нервный центр, в котором происходят первичная обработка зрительных сигналов, преобразование их в нервные импульсы, передающиеся в головной мозг. Сигналы в сетчатке передаются через цепочку из трех основных типов клеток, которые различаются по строению и функциональным свойствам: 1) фоторецепторы (палочки и колбочки); 2) биполярные клетки; 3) ганглиозные клетки (рис. 6.9). Взаимодействие между ними обеспечивается горизонтальными и амакриновыми клетками. Горизонтальные клетки осуществляют связь на уровне переключений от фоторецепторов к биполярам, амакриновые клетки - на уровне переключений от биполяров к ганглиозным клеткам. Фоторецепторы (колбочки и палочки) составляют самый внутренний слой сетчатки. В следующем клеточном слое находятся тела горизонтальных и биполярных клеток. Терминали аксонов биполярных клеток образуют синаптические контакты с дендритами ганглиозных клеток и отростками амакриновых клеток. Отростки амакриновых клеток, в свою очередь, контактируют с телами и дендритами ганглиозных клеток, а также с другими амакриновыми клетками. Внешний слой сетчатки образован телами ганглиозных клеток и волокнами, которые в последующем образуют зрительный нерв.
Оптика глаза создает на сетчатке изображение, а при помощи аккомодации это изображение становится резким. На первом этапе зрительного восприятия световая энергия трансформируется в нервное возбуждение. Этот процесс происходит в наружных члениках фоторецепторов, содержащих фотопигмент: в колбочках - йодопсин, в палочках - родопсин. Молекула фотопигмента состоит из белковой части - опсина и части, поглощающей свет - хромофора. По химическому составу хромофор является альдегидом витамина А., Поэтому недостаток витамина А в пище приводит к ухудшению зрения в сумерках (так называемая «куриная слепота»). Колбочки и палочки отличаются друг от друга по структуре и зрительным пигментам, а также по некоторым особенностям функционирования. Однако основные принципы возбуждения у них одинаковы. Наружные сегменты фоторецепторов содержат высокоэффективную многоступенчатую систему усиления сигнала о свете. Внутриклеточные регистрации от колбочек и палочек животных показали, что в темноте вдоль фоторецептора течет темновой ток, выходящий из внутреннего сегмента и входящий в наружный сегмент. Освещение приводит к блокаде этого тока. Рецепторный потенциал модулирует выделение химического медиатора в синапсе фоторецептора. Было показано, что в темноте фоторецептор непрерывно выделяет медиатор, который действует деполяризующим образом на мембраны постсинаптических отростков горизонтальных и биполярных клеток. Гиперполяризация преси-наптической мембраны при освещении приводит к замедлению выделения медиатора, что, в свою очередь, вызывает гиперполяризацию постсинаптических нейронов. Фоторецепторы связаны между собой электрическими (щелевыми) контактами. Эта связь избирательная: палочки связаны с палочками, колбочками и т.д. Благодаря электрической связи отдельных фоторецепторов сигналы, возникающие в наружном сегменте одного фоторецептора, «расплываются» по сети связанных друг с другом рецепторов.
Начиная с уровня биполярных клеток нейроны зрительной системы, дифференцируются на две группы, противоположным образом реагирующие на освещение и затемнение: клетки, возбуждающиеся при освещении и тормозящиеся при затемнении, оn - нейроны и клетки, возбуждающиеся при затемнении и тормозящиеся при освещении, - off-нейроны. Такое разделение сохраняется на всех уровнях зрительной системы, до коры включительно. Очевидно, оно составляет основу механизма для восприятия двух противоположных классов зрительных образов: светлых объектов на темном фоне (возбуждаются оn - нейроны) и темных объектов на светлом фоне (возбуждаются off-нейроны).
Горизонтальные клетки сетчатки дают реакции, сходные с ответами биполярных нейронов, длительное изменение мембранного потенциала, сохраняющееся в течение всего времени действия света данной интенсивности. Известно, что горизонтальные нейроны сетчатки генерируют два типа потенциалов: L-тип реакции состоит в гиперполяризации на свет любой длины волны и С-тип (цветовой) - знак реакции зависит от длины волны. В настоящее время предполагают, что горизонтальные клетки играют роль регуляторов синаптической передачи от фоторецепторов к биполярам. Отростки горизонтальных клеток, простираясь вдоль наружного синаптического слоя на значительные расстояния (сотни микрометров), контактируют с синапсами, связывающими фоторецепторы и биполяры, и могут передавать сигналы вдоль наружного синаптического слоя.
В передаче сигналов от биполярных клеток на ганглиозные участвуют амакриновые клетки, которые, как и горизонтальные клетки, регулируют синаптическую передачу на следующем этапе - от биполярных клеток к ганглиозным. Дендриты амакриновых клеток разветвляются во внутреннем синаптическом слое, где контактируют с отростками биполяров и дендритами ганглиозных клеток. На амакриновых клетках оканчиваются центробежные волокна, идущие из головного мозга. Амакриновые клетки управляются от биполяров через деполяризующие синапсы: увеличение деполяризующего синаптического тока вызывает деполяризацию амакриновой клетки, а уменьшение - гиперполяризацию. Горизонтально ветвящиеся отростки амакриновых клеток могут интегрировать влияния от большого числа биполярных клеток. Выходной сигнал, возникающий от такого интегрирования, влияет на активность ганглиозных клеток сетчатки. Согласно современным представлениям клетки внутреннего ядерного слоя (биполярные, горизонтальные и амакриновые) являются основным источником электроретинограммы сетчатки позвоночных животных.
Выходными элементами сетчатки являются ганглиозные клетки. Большинство ганглиозных клеток сетчатки имеют концентрические рецептивные поля: при освещении одной из зон поля ганглиозная клетка возбуждается (оn - эффект), а при ее затемнении тормозится. Взаимоотношения могут быть обратными. В соответствии с этим различают рецептивные поля с оn - центром (возбуждаются при освещении центра) и с off-центром (возбуждаются при его затемнении) (рис. 6.10).
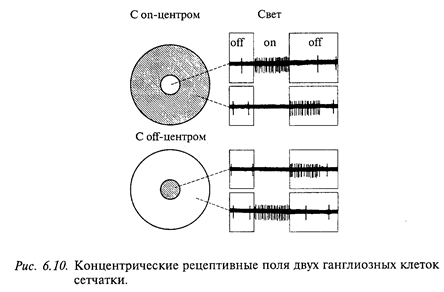
Угловые размеры центральной зоны рецептивного поля у млекопитающих варьируют от 0,5 до 8 угл. градусов (на сетчатке это соответствует пятну диаметром от 0,125 до 2 мм). Центральную зону рецептивного поля окружает концентрическая тормозная зона. Общий наружный диаметр такого концентрического рецептивного поля составляет 8-12°. Нейроны с наименьшим размером центральной зоны локализуются в центральной части сетчатки (у приматов в области fovea, у хищных, копытных и др.- в области area centralis). Тормозная кайма вокруг центральной зоны рецептивного поля не обнаруживается при низком уровне освещенности. В настоящее время предполагают, что тормозная кайма образуется горизонтальными клетками сетчатки по механизму латерального торможения, т.е. чем сильнее возбужден центр рецептивного поля, тем большее тормозное влияние он оказывает на периферию. Благодаря двум типам ганглиозных клеток с on- и off-центрами обнаружение как светлых, так и темных объектов в поле зрения обеспечивается уже на уровне сетчатки.
Как показали исследования на животных с цветовым зрением, у них существует цветооппонентная организация рецептивных полей ганглиозных клеток сетчатки, т. е. данная ганглиозная клетка получает возбуждающие и тормозные входы от колбочек, имеющих разную спектральную чувствительность (см. приложение 7). Например, если «красные» колбочки оказывают возбуждающее действие на данную ганглиозную клетку, то «синие» колбочки ее затормаживают. Обнаружены различные комбинации возбуждающих и тормозящих входов от разных классов колбочек (разных цветоприемников). Благодаря такой организации рецептивные поля отдельных ганглиозных клеток становятся избирательными к освещению определенного спектрального состава. Например, у человека, имеющего трихроматическое зрение, значительная часть цветооппонентных ганглиозных клеток связана в разных вариантах со всеми тремя цветоприемниками: возбуждение от красночувствительного приемника и торможение от сине- и зеленочувствительных приемников; возбуждение от синечувствительных и торможение от зелено- и красночувствительных и т.д.
Световая и темновая адаптация сетчатки является многокомпонентным процессом и складывается из целого ряда последовательных реакций. Самые периферические процессы -изменение диаметра зрачка и ретиномоторный эффект. Ретиномоторный эффект у позвоночных состоит в том, что от яркого света экранируются только палочки за счет движения меланина по отросткам пигментного эпителия между наружными сегментами фоторецепторов. Значительно меньше исследованы механизмы адаптации на уровне самих рецепторов и соответствующих синапсов. В них, по-видимому, принимают участие ионы кальция, а также имеет место изменение переходных процессов в горизонтальных клетках.
Анализ зрительных сигналов нейронами наружного коленчатого тела. В наружное коленчатое тело каждой стороны мозга поступают волокна от сетчаток обоих глаз. Поскольку зрительные волокна по пути к наружному коленчатому телу перекрещиваются (зрительный перекрест), к наружному коленчатому телу приходят волокна только от половины сетчатки каждого глаза: от темпоральной половины ипсилатеральной сетчатки и от назальной половины контралатеральной сетчатки (см. рис. 6.9).
В различные слои наружного коленчатого тела приходят разные ретинальные волокна. Например, у обезьяны наружное коленчатое тело состоит из шести слоев. Топическая проекция поля зрения имеется в каждом из слоев коленчатого тела. Хотя в разные слои наружного коленчатого тела приходят ретинальные волокна от разных сетчаток (контра- и ипсилатеральной), эти проекции расположены одна под другой, так что можно выделить колонкообразный участок, пересекающий все слои коленчатого тела, который соответствует проекции одной точки поля зрения. При этом наиболее подробно в коленчатом теле представлена проекция центральной части поля зрения (на сетчатке приматов соответствует fovea).
Так же как и рецептивные поля ганглиозных клеток, все нейроны наружного коленчатого тела можно разделить на два класса: с оn - центром (освещение центра рецептивного поля активирует нейрон) и с off-центром (нейрон активируется затемнением центра). Размер оn -зоны (у кошки) до 2°, off-зоны - до 5°. В наружном коленчатом теле имеется примерно равное количество нейронов с on- и off-центрами. Сходное строение рецептивных полей этих двух уровней зрительного анализатора (сетчатки и коленчатого тела) дает основание предполагать, что в структуре рецептивных полей нейронов наружного коленчатого тела отражаются свойства рецептивных полей ганглиозных клеток сетчатки.
Подобно нейронам сетчатки, нейроны наружного коленчатого тела у животных с цветовым зрением обладают цветовой чувствительностью. Так, в коленчатом теле обезьяны обнаружены нейроны, центр рецептивного поля которых связан с одним из цветоприемников, а периферия рецептивного поля - с другим. При этом реакция нейрона на стимуляцию периферии рецептивного поля всегда имеет противоположный знак реакции на стимуляцию центра. Наружное коленчатое тело - это первый уровень, на котором в зрительной системе млекопитающих вследствие неполного перекреста зрительных волокон происходит конвергенция от двух сетчаток. На схеме зрительной системы это отражено перекрытием полей зрения правого и левого глаз (см. рис. 6.8). Конвергенция является необходимым условием стереоскопического восприятия трехмерного мира.
Анализ зрительных стимулов нейронами коры больших полушарий. Каждое афферентное волокно от наружного коленчатого тела разветвляется в коре на площади в несколько сотен микрон. Аксоны некоторых звездчатых нейронов покидают поле 17 и направляются в другие области мозга. В полях 18 и 19 коры хорошо развит III слой, в котором находятся пирамидные нейроны с развитыми апикальными дендритами. В V и VI слоях много веретенообразных и треугольных клеток. Ассоциативные волокна поля 17 направляются в поля 18, 19, 21 и 7. Из полей 17, 18 и 19 эфферентные волокна направляются в переднее двухолмие и претектальную область, в подушку зрительного бугра (задняя часть таламуса), наружное коленчатое тело. Сетчатка отдельно представлена во всех трех главных зрительных полях (поля 17, 18 и 19). Наиболее упорядоченное топологическое соответствие имеет место между сетчаткой и полем 17. Представительство сетчатки, например, в поле 17 (затылочный полюс), организовано следующим образом. В задней части поля 17 локализуется центральная часть сетчатки. Продвижение по коре в каудальном направлении соответствует переходу в верхнюю часть, а в ростральном направлении - в нижнюю часть сетчатки.
В отличие от наружного коленчатого тела основная масса нейронов зрительных областей коры больших полушарий (поля 17, 18 и 19) специализирована на выделении ориентированных линий и контуров, которые составляют основные элементы зрительных стимулов. Указанная способность полностью обусловлена строением рецептивных полей корковых нейронов. В отличие от концентрических рецептивных полей предшествующих уровней зрительного анализатора (сетчатка и наружное коленчатое тело) рецептивные поля корковых нейронов имеют параллельно расположенные антагонистические зоны, которые определенным образом ориентированы в поле зрения. Рецептивное поле коркового нейрона, представленное на рис. 6.11, называется «простым». Рецептивное поле такого нейрона действует как своеобразный шаблон. Если зрительный стимул совпадает с этим шаблоном, нейрон реагирует. Избирательность реакции такого нейрона полностью определяется организацией его рецептивного поля. При одновременной стимуляции нескольких точек однородной полосы (on- или off-) этого поля ответы суммируются. Наиболее интенсивный ответ наблюдается в случае стимуляции рецептивного поля полоской (темной или светлой в зависимости от характера on- или off-рецептивного поля). Однако если стимулирующая полоска одновременно покрывает и антагонистическую зону, то ответ нейрона резко уменьшается. Благодаря такой организации рецептивного поля нейрон реагирует не на общий уровень освещенности поля зрения, а на контраст, т. е. выделяет контуры изображения.
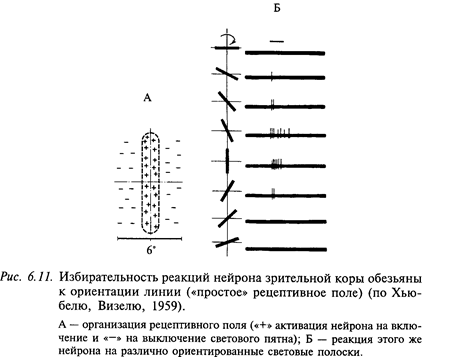
Кроме нейронов с «простыми» рецептивными полями, в зрительной коре млекопитающих описаны нейроны, избирательно чувствительные к ориентированным стимулам, но не имеющие в рецептивном поле четко выраженных антагонистических зон. Нейроны этого типа имеют слабую реакцию при стимуляции их рецептивного поля точечным стимулом, но зато хорошо реагируют на оптимально ориентированные полоски. Такая же полоска с другой (не оптимальной) ориентацией либо не вызывает реакции, либо эти реакции очень слабые. Таким образом, торможение реакции возникает при стимуляции тех же участков рецептивного поля, которые при действии оптимально ориентированного стимула ведут себя как возбуждающие. Эти нейроны имеют «сложные» рецептивные поля (рис. 6.12). Характерным свойством нейронов с такими рецептивными полями является их способность реагировать на рисунки, не содержащие определенным образом ориентированных линий, например, беспорядочно чередующиеся светлые и темные пятна нерегулярной формы. Это свойство сложных нейронов в сочетании с их относительно слабой избирательностью может указывать на то, что они приспособлены для выделения сложных зрительных стимулов. Кроме двух указанных выше типов рецептивных полей, у корковых нейронов описаны также сверхсложные рецептивные поля. Для оптимальной стимуляции сверхсложного нейрона важно, чтобы стимулирующая полоска была не только оптимально ориентирована, но и имела бы оптимальную длину. Увеличение длины полоски сверх определенной длины приводит к торможению реакции нейрона. Это происходит вследствие наличия в их рецептивном поле дополнительных «фланговых» тормозных зон на краях рецептивного поля. Рецептивное поле сверхсложного нейрона может иметь одну или две тормозные фланговые зоны. В связи с этим для максимального возбуждения нейрона необходимо ограничить длину полоски либо с одной стороны, либо с обеих. В первом случае нейрон становится чувствительным к появлению в его рецептивном поле углов. Во втором случае он максимально активируется на определенный размер стимула (его длина ограничивается расстоянием между двумя фланговыми зонами).
Основой бинокулярного стереоскопического зрения является диспаратность - оценка различия проекций изображений на сетчатках обоих глаз. Известно, что 84% нейронов зрительной коры бинокулярны. Они реагируют при одновременной стимуляции двух сетчаток, при этом нейроны одной колонки имеют близкие значения диспаратности. Благодаря тому, что правый и левый глаза рассматривают один и тот же объект с разных точек, изображения объекта на правой и левой сетчатке сдвинуты относительно друг друга. Это различие изображений зависит от того, насколько приближен или отдален объект относительно точки пересечения оптических осей двух глаз (точка фиксации). Нейрофизиологической основой бинокулярного зрения является взаимодействие двух рецептивных полей, которые имеются у каждого бинокулярного нейрона зрительной коры (на контра - и ипсилатеральной сетчатке). Каждый бинокулярный нейрон избирателен к какой-то одной определенной диспаратности (это определяется диспаратностью его рецептивных полей), а поскольку диспаратность зависит от удаленности объекта, реакции бинокулярных нейронов оказываются избирательными к определенной удаленности. В коре имеется целый набор нейронов с разной диспаратностью. Эта совокупность нейронов составляет механизм, измеряющий удаленность объекта. Описанный выше механизм диспаратности лежит в основе стереоскопического зрения.

Цветооппонентные нейроны в зрительной коре обезьян. Значительная часть цветооппонентных нейронов зрительной коры приматов имеет простые или концентрические рецептивные поля. Большинство нейронов с цветооппонентными рецептивными полями встречаются в слое IV. Для нейронов этого класса характерна цветовая оппонентность в пределах центра рецептивного поля: нейрон реагирует возбуждением на стимуляцию одного цветоприемника в центре рецептивного поля и тормозится при стимуляции другого. Одни нейроны реагируют оn - ответом на красное освещение и off-ответом - на зеленое, реакция других - обратная. Белый свет не вызывает реакции этих нейронов, так как антагонистические влияния от двух цветоприемников взаимно погашаются. У нейронов с концентрическими рецептивными полями, помимо оппонентных отношений между цветоприемниками, существуют так же обычные для концентрических полей антагонистические отношения между центром и периферией. В результате возникает структура с двойной цветооппонентностью. Если воздействие на центр рецептивного поля вызывает, например, оn – ответ на красное освещение и off – ответ – на зеленое, то нейрон с такими свойствами сочетает избирательность к цвету с избирательностью к локальным изменениям яркости пятна соответствующего цвета. Эти нейроны не реагируют ни на стимуляцию белым светом (из-за оппонентных отношений между цветоприемниками), ни на диффузную стимуляцию светом любой длиной волны (из-за антагонистических отношений между центром и периферией рецептивного поля). Максимум реакции регистрируется, если одновременно стимулировать разные цветоприемники в центре и на периферии рецептивного поля. Цветооппонентные нейроны, избирательные к ориентации, могут иметь простые и сложные рецептивные поля. В простом рецептивном поле различают две или три параллельно расположенные зоны, между которыми имеется двойная оппонентность: если центральная зона имеет оn - ответ на красное освещение и off - ответ на зеленое, то краевые зоны дают оff - ответ на красное и оn - ответ на зеленое. В сложном рецептивном поле нет раздельных зон с разной спектральной чувствительностью. Нейроны этого типа реагируют только на определенным образом ориентированные линии предпочитаемого цвета или цветоконтрастные границы. Большинство цветооппонентных нейронов коры у обезьян связано с красно- и зеленочувствительными цветоприемниками. С синечувствительными цветоприемниками связано значительно меньше нейронов. Предполагается, что информация, выделенная нейронами первичного зрительного поля 17, далее передается для обработки во вторичную (поле 18) и третичную (поле 19) области коры.
Колончатая организация зрительной коры. При погружении микроэлектрода перпендикулярно поверхности зрительной коры, как и в других корковых проекционных зонах (моторной, слуховой и т. д.), по ходу электрода всегда встречаются нейроны с близкими свойствами. Часто эти нейроны объединяются по признаку организации своих рецептивных полей: в одной колонке собраны нейроны примерно с одной ориентацией рецептивных полей. Упорядоченность в расположении ориентационных колонок очень высока в зрительной коре обезьян. Смещение регистрирующего микроэлектрода в коре на каждые 25 - 50 мкм в тангенциальном направлении приводит к повороту рецептивного поля регистрируемых нейронов в одном и том же направлении (по часовой стрелке или против) на величину примерно 10°, так что полный набор колонок со всеми ориентациями рецептивных полей в пределах 180° занимает в коре участок 500-1000 мкм. В пределах этого участка отдельные колонки в зрительной коре обезьяны в поперечном (параллельно поверхности коры) сечении имеют вид узких полосок шириной 25-50 мкм. Под гиперколонкой понимается участок коры, включающий набор ориентированных колонок (с ориентациями в пределах 180° и перекрывающиеся с ними две окулодоминантные колонки). Такая гиперколонка обрабатывает информацию от определенного участка сетчатки; информация от соседних участков обрабатывается такими же соседними гиперколонками. Однако из вышеизложенного не следует, что нейроны, составляющие колонку, в функциональном отношении совершенно однотипны. Например, в зрительной коре кошки одна колонка содержит нейроны с простыми и сложными рецептивными полями. В этом случае общим признаком будет только ориентация рецептивных полей отдельных нейронов. В одной колонке могут находиться нейроны с одинаковой цветоизбирательностью и другими однотипными свойствами. Обнаружено, что упорядоченность в размещении нейронов зрительной коры обезьян имеет место не только по вертикали в виде колонок, но и по горизонтали. Так, например, нейроны с простыми рецептивными полями находятся преимущественно в глубине III и IV слоев. Нейроны со сложными и сверхсложными рецептивными полями локализованы в основном в верхних (II и верхняя часть III) и нижних (V и VI) слоях коры.
Способен ли новорожденный видеть зрительный мир так, как его видит взрослый? Эксперименты на животных показали, что многие свойства зрительной системы заложены с рождения, но развитие этих способностей зависит в дальнейшем от воспитания. Например, было установлено, что у 1 - 2-недельных котят избирательные свойства корковых нейронов уже сформированы, т.е. они образовались исключительно за счет генетических программ в отсутствие зрительных стимулов (котята в этот период еще слепые). Вместе с тем если у котенка в течение первых 2-3 мес. жизни исключить предметное зрение, например, одним глазом, то нейроны коры, соответствующие этому глазу, теряют ориентированные зрительные поля. Интересно, что свойства нейронов наружного коленчатого тела при этом заметно не страдают. Отсюда был сделан вывод, что свойства корковых нейронов определяются исключительно внутрикорковыми связями. У котят период, чувствительный к депривации, начинается после 4-5 недель после рождения и длится до 6-8 недель. У низших обезьян этот период значительно дольше: начинается через 1-2 мес. после рождения и продолжается до 1,5-2 лет. Интересные результаты были получены в экспериментах с воспитанием котят в определенной зрительной среде. Например, если котята в течение первых месяцев после рождения видят только вертикальные (черно-белые) или только горизонтальные полосы, то в более поздние сроки в зрительной коре обнаруживаются нейроны с ориентацией рецептивных полей, соответствующих зрительной среде периода воспитания. Из этих экспериментов было сделано два важных вывода: 1) межнейронные связи, соответствующие данному виду животного, программируются генетически; 2) в то же время в раннем онтогенезе имеется период, чувствительный к воздействию внешних условий среды обитания данного вида. Такое двойное обеспечение межнейронных связей представляется биологически целесообразным.
Наблюдения на людях подтверждают выводы экспериментальных исследований. Например, после снятия катаракты, образовавшейся в раннем детском возрасте, предметное зрение остается утерянным. Это несмотря на то, что такого человека нельзя назвать полностью слепым: он различает свет и темноту. У человека формирование предметного зрения, по-видимому, занимает интервал до 15 лет.
Дата добавления: 2016-06-05; просмотров: 1868;
Поиск по сайту
Узнать еще
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- I.2. Антигены системы АВ0. Генетика. Структура
- I.2.1 ПЕРВЫЙ ЗАКОН НЬЮТОНА. ИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА.
- I.2.1 ПОЛНАЯ И ВНУТРЕННЯЯ ЭНЕРГИЯ СИСТЕМЫ. ТЕПЛОТА И РАБОТА
- I.3. Антитела системы АВ0
- I.5.4 НЕИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА. СИЛЫ ИНЕРЦИИ
- II. ЭЛЕКТРИЧЕСКИЙ ДИПОЛЬ. ДИПОЛЬНЫЙ МОМЕНТ СИСТЕМЫ ЭЛЕКТРИЧЕСКИХ ЗАРЯДОВ
- III. Избирательные системы.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
