Эпистаз— взаимодействие неаллельных генов, при котором ген одной аллельной пары подавляет действие доминантного гена другой аллельной пары.
Генотип как целостная система
Взаимодействие генов— это совместное действие нескольких генов, в результате которого появляется признак, которого нет у родителей, или усиливается проявление уже имеющегося признака.
Для объяснения результатов взаимодействия генов важно понимать механизм формирования признаков.
Ген — это участок ДНК, в котором закодирована информация об одном белке. В простейшем случае формирование признака может происходить в результате действия одного белка, синтез которого определяется одним геном:
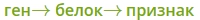
— но обычно признак формируется в результате сложных биохимических процессов. В клетке происходит взаимодействие между белками-ферментами, синтез которых определяется генами, или между веществами, которые образуются под влиянием этих ферментов.
Возможны следующие типы проявления генов в фенотипе:
· один признак формируется в результате взаимодействия нескольких белков, синтез которых определяется несколькими генами:
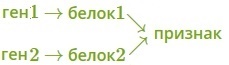
· один ген определяет синтез белка, который влияет на формирование нескольких признаков:
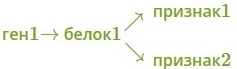
— как показано на схеме.
Возможно взаимодействие как между аллельными генами, так и между неаллельными.
Аллельные гены расположены в одинаковых участках гомологичных хромосом и определяют один признак. Примеры взаимодействия аллельных генов:
· полное доминирование;
· неполное доминирование.
(Сведения о взаимодействии аллельных генов изложены в предыдущих темах.)
Неаллельные гены расположены в разных участках негомологичных хромосом. Формы взаимодействия неаллельных генов:
· комплементарность;
· эпистаз;
· полимерия.
Комплементарность, эпистаз и полимерия — это взаимодействия, при которых несколько генов определяют один признак.
Наследование, при котором один ген влияет на формирование нескольких признаков, называется плейотропия (или множественное действие гена).
Комплементарность
Комплементарность(дополнительное действие генов) — взаимодействие неаллельных генов, при котором они дополняют друг друга, а признак формируется лишь при одновременном действии двух доминантных генов, каждый из которых в отдельности не вызывает развития признака.
Пример: наследование окраски цветков у душистого горошка:
| генотип | признак |
| A−B− | фиолетовые цветки |
| A−bb | белые цветки |
| aaB− | белые цветки |
| aabb | белые цветки |
Ни один из доминантных генов не определяет окраску цветка. Фиолетовая окраска появляется только при наличии в генотипе растения двух доминантных генов A и B.
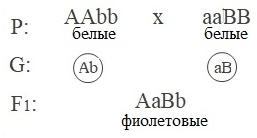
При скрещивании дигетерозигот результаты следующие.
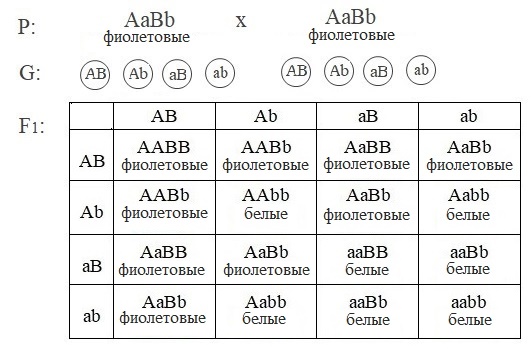
Расщепление по генотипу: 9A−B−:3A−bb:3aaB−:1aabb.
Расщепление по фенотипу: 9 (фиолетовые) :7 (белые).
При комплементарном взаимодействии генов могут наблюдаться и другие типы расщепления признака.
У людей данный тип взаимодействия обуславливает нормальное развитие слуха (один ген определяет развитие улитки, а другой ген — развитие слухового нерва). Нормальный слух формируется при наличии двух доминантных генов. Если один из генов присутствует в доминантной форме, а второй — в рецессивной, то проявляется глухота, так как один из органов слухового анализатора не развивается.
I. Комплементарность (дополнительность)
Комплементарность – взаимодействие генов, при котором доминантные аллели двух генов при совместном нахождении в генотипе (А-В-) обусловливают развитие нового фенотипа по сравнению с тем, что обусловливает каждый ген в отдельности (А-вв, ааВ-).
1. Наследование окраски цветков у душистого горошка.
| Р | ♀ ААвв | × | ♂ааВВ |
| бел | бел | ||
| F1 | ♀ АаВв | × | ♂ АаВв |
| крас | крас | ||
| F2 | 9 А-В-: (3А-вв + 3 ааВ- + 1аавв) | ||
| 9 крас | : | 7 бел |
А-– наличие пропигмента
В- – наличие фермента
2. Наследование окраски глаз у дрозофилы.
| Р | ♀ ААВВ | × | ♂аавв |
| крас | бел | ||
| F1 | ♀ АаВв | × | ♂ АаВв |
| крас | крас | ||
| F2 | 9А-В-: 3А-вв : 3 ааВ- : 1аавв | ||
| 9 крас: 3яр-кр: 3 кор :1 бел |
3. Наследование окраски шерсти у кролика.
А- – пигмент может синтезироваться.
аа – пигмент не может синтезироваться.
В- – пигмент нормальный.
вв – пигмент ослабленный.
| Р | ♀ ААвв | × | ♂ааВВ |
| гол | бел | ||
| F1 | ♀ АаВв | × | ♂ АаВв |
| черн | черн | ||
| F2 | 9 А-В- : 3А-вв : (3 ааВ- + 1аавв) | ||
| 9 черн : 3 гол : 4 бел |
4. Наследование формы плода у тыквы.
| Р | ♀ ААвв | × | ♂ ааВВ | ||
| сфер | сфер | ||||
| F1 | ♀ АаВв | × | ♂ АаВв | ||
| диск | диск | ||||
| F2 | 9А-В- : 6(3А-вв + 3 ааВ-): 1аавв | ||||
| 9 диск: | 6 сфер | :1 удлин | |||
(Комплементарность.Впервые взаимодействие неаллельных генов было обнаружено в начале 20-го столетия при анализе наследования формы гребня у кур. Разные породы кур (леггорны, виандоты, европейские, малайские) имеют соответственно листовидный, розовидный, гороховидный и ореховидный гребни.
В результате скрещивания кур, имеющих розовидный и гороховидный гребни, в потомстве первого поколения (F1)возникает новая ореховидная форма гребня (новая форма гребня, возникает из-за взаимодействия геновАиВ). Скрещивание гибридовF1приводит к следующим результатам во втором поколении(F2).
Розовидный х Гороховидный
Р 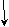 : ААвв ааВВ
: ААвв ааВВ
G АаВв
 F1: Ореховидный
F1: Ореховидный
Рис. 64.Формы гребней у кур (слева направо: розовидный, гороховидный, ореховидный, листовидный)
 ♂
♀ ♂
♀
| АВ | Ав | аВ | ав |
| АВ | Орех. ААВВ | Орех. ААВв | Орех. АаВВ | Орех. АаВв |
| Ав | Орех. ААВв | Розов. ААвв | Орех. АаВв | Розов. Аавв |
| аВ | Орех. АаВВ | Орех. АаВв | Горох. ааВВ | Горох. ааВв |
| ав | Орех. АаВв | Розов. Аавв | Горох. ааВв | Листов. аавв |
Впоследствии такой тип взаимодействия неаллельных генов, при котором совместно присутствующие в генотипе организма неаллельные гены обусловливают развитие нового признака, был назван комплементарностью.
К комплементарным относятся такие гены, которые при совместном действии в генотипе в гомо- (ААВВ) или гетерозиготном (А-В-) состоянии обусловливают развитие нового признака. Присутствие доминантных аллелей двух генов А и В у 9/16 кур второго поколения ведёт к образованию нового фенотипа - ореховидного гребня. Действие же каждого гена в отдельности (А-вв или ааВ-) ведёт к воспроизведению признака лишь одного из скрещиваемых родителей.
У человека по типу комплементарности взаимодействуют доминантные неаллельные гены MиR. Образование чёрного пигмента меланина контролирует ген М, который представлен в популяциях людей тремя аллелями:MBk, MBw, MBd. Доминантный аллель (R2) второго гена контролирует синтез красного пигмента. Сочетание аллелей названных генов обусловливает весь спектр пигментации волос у человека. Исключение составляют альбиносы, гомозиготные по гену альбинизма, который локализован в ином локусе и вызывает полное отсутствие синтеза пигмента в организме.
Эпистаз
Эпистаз— взаимодействие неаллельных генов, при котором ген одной аллельной пары подавляет действие доминантного гена другой аллельной пары.
Гены, которые подавляют действие других генов, называют генами-супрессорами, или генами-ингибиторами.
Реакция между данными генами может быть доминантной и рецессивной.
Доминантный эпистаз наблюдается в случае, когда доминантная аллель гена-супрессора (обычно он обозначается буквой I) подавляет действие другого гена.
Рецессивный эпистаз наблюдается, когда рецессивная аллель гена-супрессора угнетает проявление любой из аллелей другого гена.
Расщепление по фенотипу при каждом из видов этих взаимодействий различается. При доминантном эпистазе при скрещивании дигетерозигот расщепление может быть следующим — 13:3, 7:6:3 или 12:3:1. При рецессивном эпистазе расщепление такое: 9:3:4, 9:7 или 13:3.
Пример:
у большинства пород кур наличие пигмента в перьях определяет доминантный аллель A. Ген I подавляет синтез пигмента.
| Генотип | Признак |
| A−I− | белые перья |
| A−ii | окрашенные перья |
| aaI− | белые перья |
| aaii | белые перья |
При скрещивании двух белых дигетерозиготных птиц в потомстве появляются цыплята с окрашенным оперением.
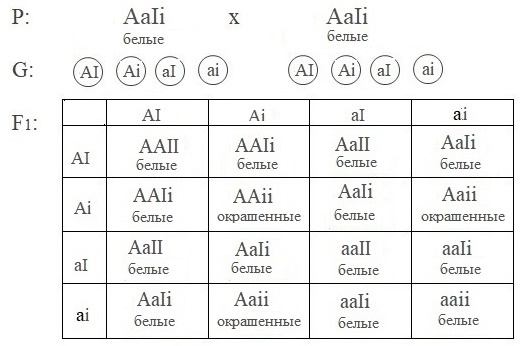
Расщепление по фенотипу: 13 (белые) :3 (окрашенные).
II. Эпистаз
Эпистаз – тип взаимодействия аллелей двух генов, при котором аллели одного гена подавляют действие аллелей другого гена.
Гены, подавляющие действие других генов, называют супрессорами, или ингибиторами.
Эпистаз бывает доминантным и рецессивным.
При доминантном эпистазе доминантный аллель одного из генов может выполнять функцию только ингибитора, подавляющего действие доминантного аллеля другого гена. Ингибитор (супрессор) обозначается буквой I или S, его рецессивный аллель, который такой функцией не обладает, – i или s. Функциональный аллель обозначают С или с. I- > С-
1. Наследование окраски луковицы у лука.
| Р | ♀ ССII | × | ♂ ccii |
| бел | бел | ||
| F1 | ♀ CcIi | × | ♂ CcIi |
| бел | бел | ||
| F2 | (9C-I- + 3 ccI- + 1 ccii) | : 3C-ii | |
| 13 бел | 3 окр |
2. Наследование окраски шерсти у собак.
| С- – черн | Р | ♀ ccii | × | ♂ ССII |
| I- – ингибитор | кор | бел | ||
| F1 | ♀ CcIi | × | ♂ CcIi | |
| бел | бел | |||
| F2 | (9C-I- + 3 ccI-) | : 3C-ii | : 1 ccii | |
| 12 бел | : 3 чер | : 1 кор |
Рецессивный эпистаз проявляется в том, что рецессивные аллели одного гена в гомозиготном состоянии подавляют действие доминантного аллеля другого гена. аа > В-
В F2 при этом проявляется 3 фенотипа в соотношении 9:3:4 (см. п. 3 – комплементарность).
(Противоположным комплементарности типом взаимодействия неаллельных генов является, по своей сути,
Полимерия
Многие количественные и некоторые качественные признаки у живых организмов определяются не одной, а несколькими парами взаимодействующих генов. В этом случае, чем больше в генотипе доминантных генов, тем сильнее проявляется признак. Такая форма взаимодействия неаллельных генов называется полимерией.
Дата добавления: 2021-11-16; просмотров: 613;
Поиск по сайту
Узнать еще
- A. Узагальнені координати і узагальнені швидкості та прискорення
- API как средство интеграции приложений.
- C04 ППВ с комментариями и примерами
- Cыры, созревающие при участии слизи.
- F00 Деменция при болезни Альцгеймера
- F50 Расстройства приема пищи
- F51 Расстройства сна неорганической природы
- F63 Расстройства привычек и влечений

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
