Динамика биологических мембран. Фазовые переходы фосфолипидных молекул.
Функционирование биомембран зависит от микровязкости липидного бислоя, от подвижности липидных молекул, от их фазового состояния. Липидная фаза мембран в физиологических условиях находится в жидком агрегатном состоянии. Молекулы жидкости, как и молекулы твердого вещества, совершают колебательные (и вращательные) движения около положения равновесия. Через определенное время (так называемое "время оседлой жизни") происходит перескок молекулы в другое положение равновесия. Время оседлой жизни τмолекул жидкости значительно меньше, чем молекул твердых веществ. В частности, для молекул фосфолипидов в составе биомемембран τ =10-7 - 10-8 с. Как уже отмечалось выше, молекулы в мембране расположены не беспорядочно, а в определенном порядке: фосфолипидные молекулы находятся в двойном слое, их гидрофобные хвосты приблизительно параллельны друг другу. Есть порядок и в ориентации гидрофильных головок липидов. Такое физическое состояние вещества, при котором агрегатное состояние жидкое, а в расположении молекул наблюдается определенный порядок, называется жидкокристаллическим состоянием. Жидкие кристаллы могут образовывать так называемые «длинные молекулы», поперечные размеры которых значительно меньше продольных. Бислойная липидная мембрана соответствует смектическому жидкокристаллическому состоянию, когда молекулы располагаются параллельно друг другу и располагаются слоями.
Коэффициент вязкости h (динамической вязкости) биологических мембран составляет 30 -100 мПа с. Такой величиной вязкости обладает подсолнечное масло при нормальных условиях. Вязкость воды при этих условиях равняется 1 мПа с. Использованием флуоресцентной, ЭПР, ЯМР – спектроскопии показана высокая подвижность фосфолипидных и белковых молекул в мембране. Подвижность этих молекул обуславливает латеральную (боковую) диффузию - хаотичное перемещение этих молекул в плоскости мембраны. При латеральной диффузии соседние молекулы скачкообразно меняются местами, и вследствие таких перескоков, молекула перемещается по мембране. Перемещение среднестатической молекулы можно оценить по коэффициенту диффузии через среднеквадратичное смещение:
D = Х־2 / 2 Δ t
Показано, что среднеквадратичное смещение фосфолипидной молекулы по мембране эритроцита составило 5 мкм/с, что сравнимо размером самого эритроцита. Таким образом, молекула липида за секунду может «обежать» клетку по периметру. Для белковых молекул среднеквадратичное смещение составляет около 0,2 мкм/с. Соответственно, коэффициенты диффузии для этих молекул составляют Dлип ≈6 ∙10-12 м2/с, Dбел ≈ 1 ∙10-14 м2/с .
Частота перескоков (число перескоков в секунду ) молекулы при латеральной диффузии рассчитывается по формуле:
v = 2 √ 3 D/ S ,
где S - площадь, занимаемая одной молекулой.
Для молекул фосфолипидов S≈7 ∙10-19 м2,соответственно, v =3 ∙107 с-1.В среднем, каждая молекула липида в составе мембраны, претерпевает несколько миллионов перескоков в секунду. Как видно, время одного перескока занимает 10-7 - 10-8 с, такое же как и время оседлой жизни липидной молекулы.
Мембранные молекулы могут также диффундировать и поперек мембраны, т.е. с одной поверхности мембраны на другую. Это тип диффузии называют флип-флоп переходом (диффузией). Диффузия молекул по этому типу происходит значительно медленнее, чем латеральная диффузия. Среднее время флип-флоп перехода одной молекулы фосфолипида составляет около одного часа, в миллиарды раз больше среднего времени молекулы при латеральной диффузии.
Жидкокристаллические структуры очень чувствительны изменению параметров: температуры, давления, химического состава, электрического поля. Этим свойством объясняется высокая динамичность липидных бислойных мембран. Даже небольшие изменения этих параметров сопровождаются структурными перестройками мембран. Наиболее изученным является изменения структуры мембран при понижении температуры, так называемые фазовые переходы. При охлаждении до определенной температуры , фосфолипидная часть мембраны переходит из жидкокристаллического в твердокристаллическое (гелеобразное) состояние. В гель-состоянии молекулы расположены еще более упорядочено, чем жидкокристаллическом состоянии. Углеводородные гидрофобные хвосты липидов располагаются строго параллельно друг другу. Толщина бислоя при таком состоянии несколько больше, чем в жидком кристалле (Рис. 4). Объем занимаемый одной молекулой липида в гель-состоянии (≈0,5 нм2) несколько ниже, чем в жидкокристаллическом (≈0,6 нм2). Температура фазового перехода мембран различных клеток может изменяться от -20 ºС до + 60 ºС. Ее значение зависит от количества ненасыщенных связей в остатках жирных кислот в молекулах липидов: чем больше число ненасыщенных связей, тем ниже температура фазового перехода.
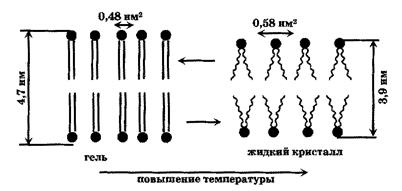
Рис. 3. Схематичное изображение структуры биологической мембраны при фазовом переходе из жидкокристаллического состояния в гель-состояние. (Антонов,26с)
Нормальное функционирование мембран осуществляется в жидкокристаллическом состоянии. Поэтому понижение температуры фазового перехода за счет изменения химического состава мембран является одной из адаптационных механизмов приспособления клеток к низким температурам. Например, значение температуры фазового перехода плазматической мембраны в клетках тканей ноги полярного оленя (от копыта до туловища) изменяется от 20 ºС до +30 ºС. Это достигается тем, что клеточные мембраны нижней части ноги содержат значительно больше ненасыщенных фосфолипидов, чем мембраны клеток в верхней части ноги оленя.
Экспериментально показано, что при фазовых переходах в липидном бислое образуются сквозные поры радиусом до 3 нм ( табл.1), которые изменяют проницаемость мембран для молекул и ионов. Увеличение проницаемости мембраны при низких температурах является одним из механизмов защиты клетки от криоповреждений. за счет выхода воды из клетки, и соответственно, снижения ее кристаллизации в цитоплазме.
Дата добавления: 2018-05-10; просмотров: 2474;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
