Рост и размножение бактерий
Рост бактерий происходит в результате множества взаимосвязанных биохимических реакций, осуществляющих синтез клеточного материала, что приводит к увеличению количества всех химических компонентов. У бактерий различают индивидуальный рост бактериальной клетки и рост бактерий в популяции.
Индивидуальный рост бактерий. О нем судят по увеличению размеров отдельных особей. Скорость роста зависит от внешних условий и физиологического состояния самой клетки. При постоянных условиях рост осуществляется с постоянной скоростью. Палочковидные бактерии растут преимущественно в направлении длинной оси, поэтому соотношение между поверхностью клетки и ее объемом при росте клеток существенно не изменяется, и это создает постоянные условия снабжения каждой части клетки питательными веществами и кислородом. Кокки растут равномерно во всех направлениях, увеличивая размеры радиуса клетки, при этом относительная величина поверхности клетки падает, поэтому условия снабжения каждой части клетки становятся все более неблагоприятными. В промежутках между клеточными делениями бактерии имеют большие размеры, чем сразу после деления.
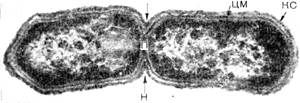
Размножение бактерий.Наиболее часто бактерии размножаются путем бинарного деления, когда из одной клетки образуется две, каждая из которых вновь делится. Процессу деления всегда предшествует репликация ДНК. Существует два типа деления – деление перетяжкой (перешнуровывание) и с помощью поперечной перегородки (рисунок 1.9).

А Б
Рисунок 1.9 – Деление бактерий
А - деление перетяжкой; Б - деление поперечной перегородкой; КС – клеточная стенка; ЦМ – цитоплазматическая мембрана; Н – нуклеоид; П – перетяжка
Деление перетяжкой (констрикция) сопровождается сужением клетки в месте ее деления, и в этом процессе принимают участие все слои клеточных оболочек. Выпячивание оболочек с обеих сторон внутрь клетки все более ее сужает и, наконец, делит на две. Так делятся многие грамотрицательным бактериям.
Деление с образованием поперечной перегородки присуще грамположительным бактериям. Однако у некоторых групп бактерий отмечена смена способов деления (тионовые бактерии, микобактерии). У шаровидных бактерий может образовываться несколько поперечных перегородок (тетракокки, сарцины).
Период от деления до деления называется клеточным циклом(онтогенез бактерий). Различают несколько типов вегетативного клеточного цикла: мономорфный – образуется только один морфологический тип клеток (например, бациллы), диморфный – два морфологических типа, полиморфный – несколько, каждый из которых характеризуется определенными и постоянными особенностями клеточного цикла (например, актиномицеты). При диморфном и полиморфном циклах различают дочерние и материнские клетки.
Почкованиеубактерий является разновидностью бинарного деления. Этот способ размножения присущ бактериям, имеющим диморфные или полиморфные клеточные циклы. Почкующимся бактериям присуща полярность клеток. Некоторые бактерии размножаются с помощью экзоспор (но не эндоспор!), фрагментами гиф (актиномицеты). Есть бактерии, у которых имеются половые ворсинки, или F-пили (англ. fertility –фертильность, плодовитость), обусловленные наличием полового фактора.
Бактерии характеризуются высокой скоростью размножения. Например, при благоприятных условиях кишечная палочка делится каждые 20…30 мин, за сутки из одной клетки проучится 272 (72 поколения). В условиях, исключающих гибель, такая биомасса составит 4720 т. Скорость размножения зависит от факторов внешней среды (температура, условия питания, влажность, реакция среды и др.) и от видовых особенностей бактерий. Высокая скорость размножения бактерий обеспечивает их сохранение на Земле даже в условиях массовой гибели. Сохранившиеся отдельные клетки размножаются и вновь дают поколение.
Рост бактерий в популяции.Популяция (фр. population – население) – это совокупность бактерий одного вида (чистая культура) или разных видов (смешанная ассоциация), развивающихся в ограниченном пространстве (например, в питательной среде). В бактериальной популяции постоянно происходит рост, размножение и отмирание клеток. Культивирование микроорганизмов в искусственных условиях бывает периодическим, непрерывным и синхронным.
Периодическое (стационарное) культивирование. Это культивирование происходит без притока и оттока питательной среды. Оно характеризуется классической кривой роста микроорганизмов, в которой выделяют отдельные фазы роста бактериальной популяции, отражающие общую закономерность роста и размножения клеток (рисунок 1.10).
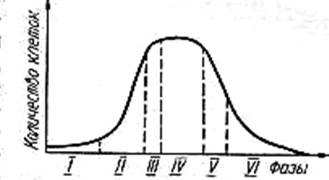
Рисунок 1.10 – Кривая роста и развития бактериальной популяции
Лаг-фаза (англ. lag – отставание) начинается с момента посева бактерий в свежую питательную среду. Клетки адаптируются к данным условиям культивирования, растут, но не размножаются, они достигают максимальной скорости роста. Абсолютная и удельная скорость роста увеличиваются от нуля до максимально возможных значений.
Абсолютная скорость роста определяется отношением:
V = dx/dt, (1.1)
где V – прирост биомассы или числа клеток;.
х – биомасса или число клеток,
t – время.
Удельная скорость роста определяется по формуле:
µ = (dx/dt) ∙ 1/х, (1.2)
где µ - прирост биомассы е единицу времени на единицу биомассы,
х – начальная биомасса.
Продолжительность лаг-фазы зависит от биологических особенностей бактерий, возраста культуры, количества посевного материала, состава питательной среды, температуры, аэрации, рН и др. Одни бактерии обладают коротким периодом задержки роста, другие длинным. Чем моложе культура, тем период короче. Чем состав питательной среды ближе к тому, в котором выращивали микроорганизмы, тем короче лаг-фаза. Изменения в питательной среде приводят к изменению лаг-фазы, так как необходимо время для синтеза ферментов, либо повышения их активности. Таким образом, факторы задержки роста можно разделить на внешние (состав среды, рН, температура и др.) и внутренние (возраст культуры). Длительность фазы моет быть от нескольких минут до нескольких часов и даже дней. В этой фазе μ = 0.
Логарифмическая, или экспоненциальная, или лог-фаза, характеризуется максимальной скоростью деления бактерий. Экспоненциальный рост популяции описывается уравнением:
Х = Хо ∙ е μmax ∙ t, (1.3)
где Хи Хо - количество клеток (или биомасса) в конце и в начале опыта;
t– время опыта;
е– основание натурального логарифма;
μmax – максимальная удельная скорость роста.
В период логарифмической фазы большинство клеток является физиологически молодыми, биохимически активными, а также наиболее чувствительными к неблагоприятным факторам внешней среды. В этой фазе μ = max.Эта фаза многостадийна, так как в начале ее бактерии растут в среде с избытком субстрата, затем концентрация его понижается, изменяется активность ферментов, возрастает содержание клеточных метаболитов. Кроме того, на рост бактерий оказывают влияние многие факторы: видовые особенности бактерий, характер питательной среды и концентрация ее отдельных компонентов, температура культивирования.
Фаза замедленного роста. Она объединяет две фазы – фазу линейного роста(μ = const) и фазу отрицательного ускорения. Фаза характеризуется в период линейного роста постоянной скоростью прироста биомассы (числа клеток). Затем при переходе в фазу отрицательного ускорения численность делящихся клеток уменьшается. Наступление фазы объясняется количественными изменениями состава питательной среды (потребление питательных веществ, накопление продуктов метаболизма).
Стационарная фаза характеризуется равновесием между погибающими и вновь образующимися клетками. Факторы, лимитирующие рост бактерий в предыдущей фазе, являются причиной возникновения стационарной фазы. Прироста биомассы нет (μ = 0).В этой фазе наблюдается максимальная величина биомассы и максимальная суммарная численность клеток. Эти максимальные величины называются урожаем, или выходом. Одним из ограничивающих факторов является максимальная концентрация клеток в единице объема питательной среды. У разных видов бактерий эта величина значительно варьирует. В стационарной фазе клетки характеризуются несбалансированным ростом (клеточные компоненты синтезируются с различной скоростью), уменьшением интенсивности обменных процессов, более высокой устойчивостью к физическим и химическим воздействиям.
Фаза отмирания (экспоненциальной гибели клеток) характеризуется уменьшением числа живых клеток, возрастанием гетерогенности популяции (появляются клетки, не воспринимающие краситель, со слабым развитием муреинового слоя и др.). Процесс отмирания превалирует над делением (μ < 0).
Фаза выживания характеризуется наличием отдельных клеток, сохранивших в течение длительного времени жизнеспособность в условиях гибели большинства клеток популяции. Выжившие клетки характеризуются низкой активностью процессов метаболизма, изменением ультраструктуры клеток (мелкозернистая цитоплазма, отсутствие полирибосом и др.). Клетки более устойчивы к неблагоприятным условиям среды.
Таким образом, при стационарном культивировании микробные клетки все время находятся в изменяющихся условиях: сначала имеются в избытке все питательные вещества, затем постепенно наступает их недостаток, затем отравление клеток продуктами метаболизма.
Влияние лимитирующих факторов на скорость роста. Для нормального роста и развития микроорганизмов среда должна содержать необходимые элементы питания, иметь соответствующую рН, температуру и т.д. Факторы, ограничивающие рост культуры, называются лимитирующими. Характерная особенность роста популяции микроорганизмов – зависимость удельной скорости роста от концентрации субстрата. Эта зависимость выражается уравнением Моно, представляющим собой гиперболическую функцию:
μ = μmax ∙ S/(S + KS), (1.4)
где μ – удельная скорость роста;
μmax- максимальная удельная скорость роста;
S – концентрация субстрата;
KS - константанасыщения, численно равная такой концентрации субстрата, которая обеспечивает скорость роста, соответствующую половине значенияμmax.
По мере потребления питательных веществ среда обогащается продуктами обмена, которые также лимитируют рост культуры. Наиболее общий случай влияния концентрации субстрата и продуктов обмена на скорость роста популяции микроорганизмов нашел отражение в модели Н.Д.Иерусалимского:
μ = μmax ∙ S/(S + KS) ∙ КР / (КР/ + Р), (1.5)
где Р – концентрация продуктов обмена;
КР- константа, численно равная такой концентрации продуктов обмена, при которой скорость роста замедляется вдвое.
Анализ этого уравнения показывает, что при условии КР >> Р, когда величиной Р можно пренебречь. скорость роста ограничена только концентрацией субстрата. Если S >> KS , то скорость роста лимитирована накоплением продуктов обмена
Непрерывное культивирование. Если в емкость, где находится бактериальная популяция, непрерывно подавать свежую питательную среду и одновременно с такой же скоростью выводить культуральную жидкость, содержащую бактериальные клетки и продукты метаболизма, то получается непрерывное культивирование. Регулируя скорость проточной среды, можно управлять ростом бактериальной популяции, например, удлинять логарифмическую или стационарную фазу на любое необходимое время. Непрерывное культивирование осуществляется в специальных приборах - хемостатах и турбидостатах.
Хемостаты. Рост бактерий регулируется концентрацией субстрата. Поддерживая постоянную концентрацию одного из необходимых субстратов (источник азота или углерода), путем регулирования скорости протока среды, можно сбалансировать скорость роста культуры. Скорость изменения величины биомассы клеток в хемостате равна разности между скоростью прироста биомассы и скоростью выноса ее из культиватора. Плотность популяции остается постоянной, если μ=D (удельная скорость роста равна коэффициенту разбавления), т.е. потеря клеток в результате вымывания и прирост их в результате размножения уравновешивается.
Турбидостаты.Принцип работы основан на регулировании скорости потока среды плотностью популяции. Плотность популяции контролируется фотоэлементом, соединенным с реле, регулирующим подачу среды. Когда плотность популяции достигает заданного уровня, реле срабатывает и в культиватор поступает свежая среде. В результате концентрация клеток уменьшается до определенного уровня и затем автоматически отключается подача среды.
Турбидостатный контроль может быть основан на других метолах определения биомассы, либо продуктов, образующихся в процессе роста бактерий (например, рН-статный способ управления скорости потока, использование оксистата– управление скоростью потока по скорости потребления кислорода и др.).
Непрерывное культивирование микроорганизмов используется для изучения их физиологии, биохимии, генетики и др., а также широко используется в микробиологической промышленности.
Синхронное культивирование. Синхронные культуры – это культуры, в которых некоторое время все клетки делятся одновременно (синхронно) за счет одинаковой готовности к делению всех особей. Синхронизация достигается физическими и химико-биологическими методами. Физические методы - это температурное воздействие, дифференциальное центрифугирование или дифференциальное фильтрование и др. Химико-биологические методы: вынужденное голодание бактерий, выращивание бактерий на неполноценных средах с последующим переносом их в полноценные среды. Синхронные культуры используются для генетических и цитологических исследований, для изучения синтеза отдельных клеточных компонентов в процессе деления бактерий.
Дата добавления: 2017-11-21; просмотров: 4652;
Поиск по сайту
Узнать еще
- Arthropoda. Паукообразные. Систематика. Географическое распространение. Морфология. Скорпионы. Пауки. Медицинское значение.
- Arthropoda.Систематика.Блохи.Виды блох.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское и эпидемиологическое значение.Меры борьбы.
- Arthropoda.Систематика.Мошки,мокрецы,слепни,оводы.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское значение,меры борьбы.
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- I.1.3 СКОРОСТЬ. УСКОРЕНИЕ.
- I.5.3 КОСМИЧЕСКИЕ СКОРОСТИ
- I.6.2 ГИДРОАЭРОСТАТИКА
- II. Абсолютное пространство по самой своей сущности, безотносительно к чему бы то ни было внешнему, остается всегда одинаковым и неподвижным».

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
