Химический состав бактерий
Химический состав бактериальной клетки сходен с химическим составом других живых организмов. Ее компонентами являются вода, минеральные и органические соединения – белки, нуклеиновые кислоты, углеводы, липиды.
Клеточная водасоставляет 75…90% массы вегетативной клетки. Нормальный метаболизм и размножение микроорганизмов возможны только в водной среде. Вода является растворителем органических и минеральных веществ, дисперсионной средой для коллоидов, источником водородных и гидроксильных ионов, а также водорода и кислорода. В клетке вода находится в двух состояниях: свободная вода, которая является растворителем, принимает участие в процессах метаболизма; связанная вода - связана с клеточными колодами, обуславливает изменение значений криоскопической точки цитоплазмы (определяет устойчивость микроорганизмов к низким и высоким температурам).
Минеральный состав. Из множества известных в настоящее время химических элементов в состав живого вещества чаще всего входит только 22, из них лишь 16 элементов встречаются во всех группах организмов. Шесть основных элементов (углерод, кислород, азот, водород, фосфор и сера) составляют 95% сухой биомассы бактериальной клетки. Углерод, азот, водород, кислород легко образуют прочные ковалентные связи посредством спаривания электронов, благодаря чему они способны давать разнообразные химические соединения. Бактерии также содержат калий, натрий, кальций, магний, хлор, железо (в пределах 1… 0,5% сухого вещества). На остальные элементы приходится около 0,3% (таблица.1.1).
Микроорганизмы используют металлы в форме катионов неорганических солей. Минеральные соли составляют от 2 до 30% сухого вещества клетки. Содержание солей варьирует в зависимости от состава питательной среды и возраста культуры. В молодых клетках их в 6…7 раз больше, чем в старых.
Таблица 1. 1 - Приблизительный элементарный состав бактериальных клеток
| Элементы | Количество, % СВ клетки | Элементы | Количество, % СВ клетки | Элементы | Количество, % СВ клетки |
| Углерод | Фосфор | Кальций | 0,5 | ||
| Кислород | Сера | Магний | 0,5 | ||
| Азот | Калий | Хлор | 0,5 | ||
| Водород | Натрий | Железо | 0,2 |
Некоторым бактериям для роста необходимы редкие элементы. По значению элементы можно условно разделить на существенные(отсутствие которых нарушает рост и размножение бактерий) и несущественные, которые могут быть заменены. Имеется группа олигоэлементов (греч. oligos - малый), которые в очень малых количествах стимулируют рост и развитие бактерий. Их общая масса не превышает 0,1% клеточного вещества. К ним относятся кадмий, ванадий, молибден, олово, никель, серебро и др. Эти элементы в питательные среды обычно не добавляют, так как они содержатся в водопроводной воде, пептоне, мясном экстракте и т.д.
Органические соединения. Большинство элементов находится в бактериальной клетке в составе биомолекул, среди которых аминокислоты, азотистые основания, сахара, жирные кислоты, спирты и т.д. являются основными компонентами органических веществ.
Белки составляют 40…80% клетки и представлены простыми и сложными белками. К сложным белкам относятся нуклеопротеиды, липопротеиды, гликопротеиды, фосфопротеиды, металлопротеиды (содержащие железо, цинк, медь), флавопротеиды. Содержание белков в бактериальной клетке варьирует в зависимости от вида бактерий, возраста культуры, состава питательной среды и т.д.
Бактериальные белки состоят из тех же 20 важнейших аминокислот, что и белки растений и животных, которые образуют полипептидные цепи (-СО-NН-).. Аминокислотный состав белков различных видов бактерий качественно и количественно различен. Только у бактерий встречаются такие аминокислоты, как диаминопимелиновая (ДАП), аминомасляная, гомосерин, D-аланин, D-глутаминовая, отсутствующие у высших организмов. Большинство бактерий сами синтезируют все необходимые им аминокислоты, но некоторые нуждаются в готовых аминокислотах, которые вносятся в питательные среды.
У бактерий преобладают глобулярныебелки, полипептидные цепи которых плотно свернуты в компактные сферические и глобулярные структуры. Большинство из них растворимы в водных растворах, легко диффундируют, для них характерны биологические функции (это ферменты, токсины и др.). Белки бактерий часто выполняют одновременно структурную и функциональную роль (например, белки цитоплазматической мембраны).
Нуклеиновые кислоты – сложные полимеры, состоящие из большого количества (от 1,5 тыс до 5 млн) мононуклеотидов. Мононуклеотиды построены из азотистого основания (пуринового – аденин, гуанин, или пиримидинового – цитозин, тимин или урацил), сахара (рибозы или дезоксирибозы) и остатка фосфорной кислоты. Мононуклеотиды ковалентно связываются между собой фосфодиэфирными связями, образуя полинуклеотиды – нуклеиновые кислоты .
Рибонуклеиновая кислота (РНК)составляет 10% сухого вещества, она обычно одноцепочная. В клетках существует три типа РНК: информационная, или матричная (используется в качестве матрицы, определяющей последовательность аминокислот в растущей полипептидной цепи); транспортная (переносит на рибосому определенные аминокислоты); рибосомальная (находится в рибосомах).
Дезоксирибонуклеиновая кислота (ДНК) составляет 3…4% сухого вещества, она состоит из двух полинуклеотидных цепей, образующих одну двойную спираль. Нуклеоид бактерий представлен гигантской молекулой ДНК, в которой закодирована вся генетическая информация клетки. Кроме того, у некоторых бактерий в цитоплазме находятся молекулы внехромосомной ДНК – плазмиды.
Углеводы в бактериальной клетке составляют 12…30% сухого вещества. Они представлены моно- и полисахаридами. Полисахаридыбактерий чрезвычайно разнообразная группа биополимеров. Среди них есть соединения, характерные, как для прокариот, так и эукариот (целлюлоза, гликоген). У бактерий обнаружен ряд полисахаридов, не встречающиеся у других организмов (тейхоевые кислоты, пептидогликаны, декстран и др.). Высокая активность полисахаридов обусловлена их способностью легко вступать в реакции с другими макромолекулами путем гидрофобных взаимодействий, а также соединяясь ионными и водородными связями. Большинство полисахаридов включает самые распространенные сахара – глюкозу, галактозу, рамнозу.
Тейхоевые кислоты – это полимеры глицерина (трехатомного спирта) или рибита (пятиатомного спирта) – глицеротейхоевая и рибитотейхоевая кислоты. В этих кислотах от 8 до 50 остатков спирта связано фосфодиэфирными связями и могут содержать моносахарид в качестве заместителя. Они содержатся в клеточной стенке бактерий (греч teichos – стенка).
Липополисахариды (ЛПС) состоят из липидной части и полисахаридной, которая представлена различными углеводами.
По функциональной активности полисахариды бактерий подразделяются на внутриклеточные (запасные вещества клетки, эндотоксины и пр.) и внеклеточные, которые входят в состав капсулы.
Липиды бактерий представлены нейтральными жирами, фосфолипидами, восками. Общее содержание липидов варьирует от 5% до 40%. Основная масса липидов в бактериальной клетке связана с другими компонентами (белками, полисахаридами). Липиды бактерий значительно разнообразнее липидов высших организмов. Они выполняют разнообразные функции – являются аккумуляторами энергии у некоторых бактерий (поли-β-оксимасляная кислота), служат структурными компонентами клетки (цитоплазматическая мембрана), участвуют в метаболизме углеводов, в энергетическом обмене, входят в состав антигенов, определяют кислотоустойчивость бактерий.
Нейтральные жиры (ацилглицерины, или глицериды) бактерий часто содержат насыщенные жирные кислоты. Ненасыщенные жирные кислоты представлены исключительно кислотами с одной двойной связью. Содержание свободных жирных кислот в клетках варьирует от 5 до 30% в зависимости от видовых особенностей бактерий и условий их выращивания. Качественный и количественный состав жирных кислот изменяется с возрастом культуры, характеризуется высокой чувствительностью к физическим и химическим факторам внешней среды. Так, молодые клетки содержат значительное количество ненасыщенных жирных кислот. Бактерии также способны включать в себя жирные кислоты из окружающей среды. Бактерии не содержат полиненасыщенные жирные кислоты, стерины и стероиды.
Фосфолипиды – основные представители бактериальных липидов. В бактериальной клетке их количество составляет от 0,4% до 6,5%. Основная масса фосфолипидов содержится в цитоплазматической мембране или в клеточных оболочках.
Воски содержат кислотоустойчивые бактерии, например, микобактерии.
Пигменты бактерий. Среди бактерий есть значительное число пигментообразующих видов. Пигменты вырабатываются в зависимости от условий выращивания культуры бактерий – минерального состава и реакции среды, источника углерода, температуры, количества кислорода, наличия света. Важными элементами для образования пигментов являются азот, магний, железо, кальций и др. Так, магний необходим для синтеза бактериохлорофиллов, железо – для продигиозина, пиоцианина. Пигменты могут выполнять в бактериальной клетке различные функции. Установлено их участие в фотосинтезе, в дыхании, в окислительно-восстановительных реакциях, защите от экстремальных факторов внешней среды (УФ-излучение, повышенная концентрация минеральных солей и т.д.).
По растворимости микробные пигменты можно разделить на две группы: нерастворимые (хромофорные) пигменты, связанные с клеточными компонентами и обуславливающие окраску колоний бактерий, но не среды; растворимые (хромоларные) пигменты, которые растворяются в питательной среде и окрашивают ее при росте бактерий. По химическому составу пигменты разнообразны – каротиноиды, меланины, хиноны, бактериохлорофиллы, пирролы.
Бактериохлорофиллами называются хлорофиллы, содержащиеся в клетках фотосинтезирующих бактерий. Их существует несколько типов, и они структурно отличаются как между собой, так и от хлорофилла высших растений. Большинство фотосинтезирующих бактерий содержит бактериохлорофилл а, у пурпурных бактерий обнаружен бактериохлорофилл b, у зеленых серобактерий – c,d,e.
Бактериохлорофиллы поглощают свет в более длинноволновой области (800…1100 нм), чем хлорофиллы высших растений.
Каротиноидные пигменты содержатся у всех фотосинтезирующие микроорганизмы и у ряда нефотосинтезирующих. Известно свыше 300 каротиноидов и они в определенной степени определяют окраску колоний.
Меланиновые пигменты (черного и коричневого цвета) – это тесно связанные с белками клетки биополимеры, которые обуславливают защиту клетки от экстремальных окислительных и восстановительных условий.
Строение бактерий
Схема строения бактериальной клетки представлена на рисунке 1.2.
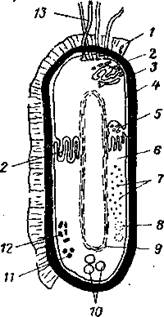
Рисунок 1.2 – Схема строения бактериальной клетки
1 – слизистая капсула, 2 – тилакоиды, 3 – клеточная стенка, 4 – цитоплазматическая мембрана, 5 – мезосома, 6 – цитоплазма, 7 – рибосомы, 8 – полисахаридные гранулы, 9 – нуклеоид, 10 – включения серы, 11 – жировые капли, 12 – гранулы полифосфатов, 13 – жгутики.
Оболочки бактериальной клетки.Цитоплазма большинства бактерий окружена оболочками: клеточной стенкой, цитоплазматической мембраной и капсульным (слизистым) слоем. Эти структуры принимают участие в обмене веществ, через оболочки клетки поступают продукты питания и удаляются продукты метаболизма. Они защищают клетку от действия вредных факторов среды, в значительной степени обуславливают поверхностные свойства клетки (поверхностное натяжение, электрический заряд, осмотическое состояние и др.). В живой бактериальной клетке эти структуры находятся в постоянном функциональном взаимодействии.
Клеточная стенка. Бактериальная клетка отделена от внешней среды клеточной стенкой. Ее толщина 10…20 нм, масса достигает 20…50% массы клетки. Это сложная полифункциональная система, определяющая постоянство формы клетки, ее поверхностный заряд, анатомическую целостность, способность к адсорбции фагов, участие в реакциях иммунитета, контакт с внешней средой и защиту от неблагоприятных внешних воздействий.
Клеточная стенка обладает эластичностью и достаточной прочностью, выдерживает внутриклеточное давление 1…2 МПа. Она имеет мельчайшие поры, через которые транспортируются продукты метаболизма.
Основными компонентами клеточной стенки являются пептидогликаны (гликопептиды, мукопептиды, муреины, гликозаминопептиды), которые содержатся только у прокариот. Специфический гетерополимер пептидогликан состоит из чередующихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных между собой посредством β-1-4-гликозидных связей, диаминопимелиновой кислоты (ДАП), D-глутаминовой кислоты, L- и D-аланина в соотношении 1:1:1:1:2. (рисунок 1.3 а).
Молекула N-ацетилглюкозамина – производное глюкозы, в котором гидроксильная группа при втором атоме углерода замещена аминогруппой (к последней присоединен ацильный остаток); N-ацетилмурамовая кислота представляет собой эфир N-ацетилглюкозамина и молочной кислоты, к карбоксильной группе которой присоединен пептидный хвост из 4…5 аминокислот. Это чаше всего L- и D-аланины, D-глутаминовая кислота и одна из аминокислот (диаминопимелиновая, диаминомасляная, лизин, L-орнитин).
Гликозидные и пептидные связи, которые объединяют субъединицы пептидогликанов, придают им структуру молекулярной сети или мешка (рисунок 1.3 б).
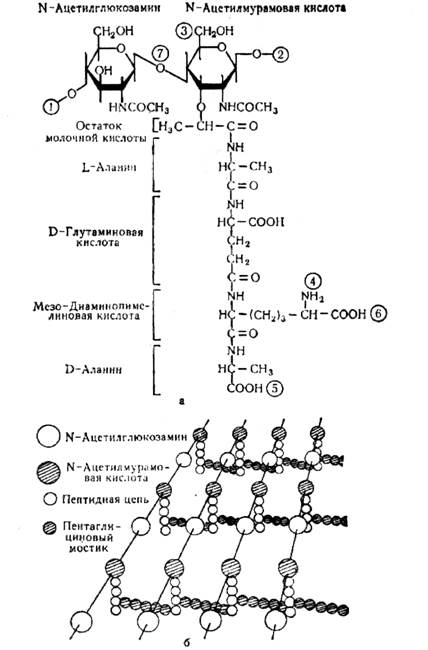
Рисунок 1.3 – Схема общей структуры пептидогликана
Окраска по Граму. Большинство бактерий в зависимости от химического состава делятся на две группы. Это свойство было впервые замечено в 1884 г. датским физиком Х. Грамом. Сущность состоит в том, что при окрашивании бактерий генцианвиалетом (кристаллвиолетом, метилвиолетом и др.) у одних бактерий краска с йодом образует соединение, которое удерживается клетками при обработке их спиртом. Такие бактерии окрашены в сине-фиолетовый цвет и получили название грамположительных (Гр+), обесцвеченные бактерии – грамотрицательных (Гр-), их докрашивают контрастной краской (фуксином). Окраска по Граму является диагностической только в отношении прокариот, обладающих клеточной стенкой. По структуре и химическому составу грамположительные и грамотрицательные бактерии существенно отличаются . их сравнительный химический состав представлен в таблице 1.2.
Грамположительные бактерии. У них клеточная стенка более толстая (20…80 нм, иногда до 100 нм), гомогенная, аморфная, содержит 50…80% (иногда до 95%) муреина, который связан с тейхоевыми кислотами.
Грамотрицательных бактерий клеточная стенка более тонкая, слоистая (внутренний ригидный слой толщиной 2…3 нм и наружный, именуемый внешней мембраной, толщиной 8…10 нм), мало муреина (5…10%) и он находится в ригидном слое, тейхоевые кислоты отсутствуют.
Таблица 1.2 - Химический состав клеточной стенки бактерий
| Компоненты клеточной стенки | Гр+ | Гр- |
| Пептидогликан (муреин) | +++ | + |
| Тейхоевые кислоты | + | - |
| Липиды | + | +++ |
| Полисахариды | + | + |
| Белки | + или - | - |
| Липополисахариды | - | + |
| Липопротеиды | - | + |
Периплазма – это пространство между внешней мембраной клеточной стенки и цитоплазматической мембраной, характерна только для грамотрицательных бактерий. Толщина ее около 10 нм, содержит муреин, белки, олигосахариды, неорганические вещества.
Цитоплазматическая мембрана (ЦПМ). К внутренней стороне поверхности клеточной стенки прилегает ЦПМ. Она составляет 8…10% массы клетки. Толщина ее равна 7…10 нм. Основным компонентом ЦПМ является липидно-белковый комплекс, в котором белки составляют 50…75%, липиды – 15…45%, могут содержаться углеводы (до 6%).
Липиды в основном представлены фосфолипидами (до 80% всех клеточных липидов). ЦПМ состоит из двойного слоя фосфолипидов. Их полярные гидрофильные головки расположены на обеих внешних поверхностях двойного слоя, а гидрофобные цепочки жирных кислот размещаются в центральной части мембраны.
Главная функция липидов – поддержание механической стабильности мембраны и придание ей гидрофобных свойств. Мембранные белки ассиметрично включены в двойной фосфолипидный слой и частично или полностью погружены в него (группа интегральных белков пронизывает насквозь слой фосфолипидов, группа периферических белков образует скопления на поверхности).
На внешней поверхности ЦПМ содержатся в основном гидролитические ферменты, на внутренней – окислительно-восстановительные. Ферменты принимают участие в окислительных процессах и в синтезе белка. ЦПМ выполняет функции внутреннего осмотического барьера, транспортные функции, синтеза веществ (мембранных липидов, компонентов клеточной стенки и капсулы и др.), энергетическую и дыхательную функции.
Мембранные образования. Скорость роста ЦПМ превышает скорость роста клеточной стенки, вследствие чего происходит инвагинация(выпячивание) ЦПМ, и возникают различной сложности внутриклеточные мембранные образования. У грамотрицательных бактерий мембранные образования развиты слабо, устроены просто, у грамположительных - хорошо развиты, более сложно организованы и образуют внутриклеточные мембранные структуры – мезосомы, тилакоиды, хроматофоры..
Капсула. Клеточная стенка снаружи окружена слизистым слоем различной толщины и конфигурации. Если слизистый слой достаточно толст, прочен и имеет определенную форму, то называется капсулой. Анатомически различают: микрокапсулы (до 0,2 мкм, неразличимы с помощью оптического микроскопа), макрокапсулы (более 0,2 мкм, видны в микроскопе), слизистый слой (вязкие, накапливающиеся на поверхности клетки вещества, иногда во много раз превышающие толщиной размеры бактериальной клетки).
По химическому составу различаются: капсулы: 1) полисахаридной природы, состоящие из гомополисахаридов (построены из сахарного остатка одного типа) или гетерополисахаридов (построены из остатков различных сахаров); 2) состоящие из полипептидов и полисахаридов. Способность образовывать капсулу является генетической, но зависит от условий среды. Капсулообразование – функция приспособления (предохраняет от высыхания, ядовитых веществ и пр.). Микроорганизмы, образующие большое количество аморфной внеклеточной слизи, часто объединяют под общим названием – слизеобразующие.
Чехлы. Отдельные виды бактерий, например, нитчатые, образуют чехлы. В отличие от капсулы чехлы имеют тонкую структуру, иногда многослойные, инкрустированные окислами металлов (железа, марганца), вследствие чего приобретают особую прочность, характеризуются более сложным составом. На дне озер и болот происходит отложение руды, состоящей из остатков железобактерий.
Придатки бактериальной клетки. На поверхности бактериальной клетки имеются придатки – ворсинки, стебельки, шипы, жгутики и др., объединенные под общим названием – пили. Строение и функции их различны. У одной и той же бактерии могут присутствовать пили разной природы. Придатки не являются жизненно важной структурой клетки, их потеря не сопровождается нарушением процесса роста и размножения бактерий.
Цитоплазма – это содержимое клеток, окруженное ЦПМ. Цитоплазма является средой, связывающей все внутриклеточные структуры в единую систему. Она представляет собой цитозоль – полужидкую коллоидную массу, состоящую на 70…80% из воды. Содержит также РНК, ферменты, продукты и субстраты метаболических реакций. Вязкость цитоплазмы примерно в 800 раз больше вязкости воды. В молодых клетках вязкость невелика, но с возрастом повышается, что является одним из факторов снижения их физиологической активности.
Нуклеоид. Электронно-микроскопические и генетические исследования установили, что бактерии имеют структуры, функционально тождественные ядрам клеток высших организмов, но отличающиеся рядом особенностей.
Нуклеоид бактерий не имеет ядерной оболочки и ДНК находится в непосредственном контакте с цитоплазмой; нет разделения на хромосомы, нить ДНК представляет собой аналог хромосомы высших организмов и называется бактериальной хромосомой (в клетке может быть несколько ее копий); отсутствует митоз и мейоз.
Рибосомы. В цитоплазме находятся мелкие зерна рибонуклеопротеидов, получивших название рибосомы, которые являются центром синтеза белка. Это частицы размером 16…18 нм, состоящие из РНК (60…65%) и белка (35…40%). Рибосомы содержат 80…85% всей РНК бактериальной клетки. В зависимости от условий и возраста в клетке содержится 1,5…50 тыс. рибосом. Скорость роста клетки определяется скоростью образования рибосом. В среднем за каждую секунду образуется около 5…10 рибосом. Большая часть рибосом объединяется в полисомы – агрегаты, состоящие из рибосом, молекул иРНК и тРНК, которые располагаются в цитоплазме, связаны с ЦПМ или мембранными структурами.
Запасные вещества образуются в клетке в результате обмена веществ, их образование зависит от условий культивирования. По консистенции запасные вещества можно разделить на жидкие (поли-β-оксибутират), полужидкие (сера) и твердые (гликоген). С химической точки зрения они могут быть химически чистыми веществами (сера) и смесями.
Безазотистые органические запасные вещества являются внутриклеточными источниками углерода и энергии. К ним относятся крахмал, гранулёза, гликоген т др. Каждый вид микроорганизма образует, как правило, только один тип запасного вещества.
Гранулёза является специфическим запасным углеводным веществом бактерий рола Clostridiun (C.butyricum, C.pasteurianum), при голодании исчезает, выявляется качественной реакцией со слабым раствором йода (клетки серо-синего или темно-синего цвета).
Гликоген – гранулы полисахарида (С6H10O4)n сферической формы, диаметром 20…100 нм, выявляются раствором йода (гранулы приобретают красно-коричневый цвет).
Углеводородные гранулы диаметром 200…300 нм, окружены белковой оболочкой, содержат углеводы того же типа, что и в среде. Встречаются у использующих углеводороды бактерий.
Поли-β-оксибутират – гранулы диаметром 00…1000 нм, окруженные однослойной белковой мембраной, окрашиваются суданом в черный или красный цвет, обнаружены только у прокариот.
Полифосфаты (волютин, или метахроматиновые гранулы) – это внутриклеточный резерв фосфора, служат источником энергии для клетки.. Эти включения называются метахроматином (греч. meta - изменение, chroma – цвет), потому что некоторые красители (например, метиленовый синий) при окрашивании гранул изменяют цвет (в фиолетово-красный), или волютином по названию бактерий Spirillum volutans.
Включения серы встречаются у серобактерий, окисляющих сероводород. Сера накапливается, когда в среде содержится сероводород, и окисляется до сульфата, когда весь сероводород среды исчерпан. Включениякарбоната кальция (известковые тельца) в клетках некоторых серобактерий выполняют функцию нейтрализаторов среды. Газовые вакуоли (или аэросомы) характерны для водных бактерий. Они являются регуляторами плавучести (снижают плотность).
Спорообразование
Бактериальные эндоспоры (греч. spora – семя) – уникальные по структуре и свойствам образования, характеризующиеся высокой устойчивостью к неблагоприятным факторам внешней среды. Они не похожи на споры других микроорганизмов, тем более на споры растений. Спорообразование не является способом размножения. Бактериальные эндоспоры – это одна из стадий развития бактерий, выработанная в процессе длительной эволюции в борьбе за сохранение вида. Из одной вегетативной клетки формируется только одна спора.
Способность к спорообразованию предопределена генетически. Но споры образуются в условиях, неблагоприятных для роста и размножения бактерий (неблагоприятные температурные условия, высушивание, недостаток питательных веществ, изменение рН среды и т.д.).
В клетке имеется два набора генов – один кодирует вегетативный рост, другой – спорообразование (эта группа генов называется спорулон). Бактериальная клетка, в которой формируется спора, называется спорангием. Процесс спорообразования составляет от полутора часов до суток, а иногда и более. Спорообразование (споруляция) происходит как в естественных условиях, так и при выращивании на питательных средах.
Спорообразование начинается с прекращения роста клетки. Изменяется направленность метаболических процессов – происходит перестройка белков клетки, часть из них синтезируется заново из внутреннего фонда свободных аминокислот; расходуются имеющиеся запасные вещества.
Стадии образования эндоспоры. Спорообразование происходит в несколько стадий (рисунок 1.4):
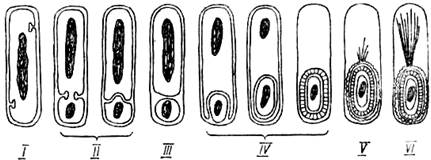
Рисунок 1.4 – Схема стадий образования эндоспоры
1. Нуклеоид вегетативной клетки приобретает компактную палочковидную форму, происходит перестройка белков.
2. Отделяется полярный нуклеоид. В результате инвагинации ЦПМ часть протопласта отделяется от материнской клетки, образуется споруляционная перегородка, которая в дальнейшем деформируется и превращается в одну из оболочек споры, появляется протопласт споры.
3. Протопласт споры окружается протопластом материнской клетки и образуется округлая проспора, окруженная двумя мембранами – внутренней и внешней, обращенной своим наружным слоем внутрь клетки. Проспора отделяется от мембраны материнской клетки и либо остается у полюса клетки, либо перемещается к ее центру. Из материнской клетки в проспору поступают некоторые аминокислоты, дипиколиновая кислота (С7Н5О4) – она отсутствует в вегетативной клетке, ионы кальция; образуется комплекс Са2+ с дипиколиновой кислотой. На второй и третьей стадиях начинают синтезироваться белки покровов споры.
4. Между двумя мембранами проспоры начинается образование муреинового слоя, которое завершается образованием толстого слоя кортекса(или коры).
5. Происходит образование покровов споры, электронно-плотной наружной оболочки споры. Все внутренние оболочки споры имеют гладкие поверхности, а наружная (или внешняя) оболочка характеризуется неровной поверхностью. Она определяет своеобразие поверхности спор разных видов бактерий. Число и строение слоистых покровов у разных видов бактерий различно.
6. Заканчивается формирование споры, появляется ее уникальное свойство – термоустойчивость. Зрелая спора имеет характерную для каждого вида бактерий форму, размеры, занимает соответствующее положение в клетке. Затем происходит освобождение споры от остатков материнской клетки (путем гибели и лизиса материнской клетки).
Положение споры в клетке. Споры могут занимать разное положение в клетке (рисунок 1.5):
- бациллярное, когда спора локализуется в клетке центрально или эксцентрально или терминально и при этом клетка не изменяет свою форму (характерно для аэробных бактерий рода Bacillus);
-клостридиальное, когда при формировании споры клетка приобретает вид веретена – эндоспора располагается в утолщенной части клетки центрально или эксцентрально;
- плектридиальное, когда спора локализуется терминально, в месте ее расположения клетка расширяется и приобретает вид барабанной палочки или ракетки. Клостридиальный и плектридиальный типы расположения эндоспор свойствен анаэробным бактериям рода Clostridium.
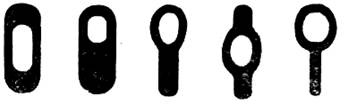
Рисунок 1.5 – Характер расположения эндоспоры в клетке
Строение зрелой споры. Строение зрелой споры разных видов бактерий однотипно (рисунок 1.6). Спороплазма (или сердцевина) содержит нуклеиновые кислоты и белки (суммарно до 50…60% сухого вещества споры), дипиколиновую кислоту (5…25%), ионы Са2+ (до 2%), ферменты (большинство из них не отличается от ферментов вегетативной клетки, но они неактивны), липиды и др. вещества, содержит также одну или несколько копий хромосом и рибосомы. Дипиколинат кальция заполняет пространство между макромолекулами спороплазмы, препятствуя их взаимодействию.
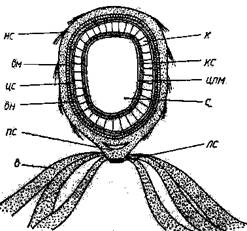
Рисунок 1.6 – Схема строения эндоспоры
с – спороплазма; цпм –цитоплазматическая мембрана; кс – клеточная стенка; к – кортекс: вн – внутренний слой споровой оболочки; нс – наружный слой споровой оболочки; цс – слой цитоплазмы между внешней мембраной споры и оболочкой; вм – внешняя мембрана споры; пс - подушечка споры; лс – линзовая структура, состоящая из гранулярного вещества; в – выросты споры
Термоустойчивость спор. Эндоспоры характеризуются высокой термоустойчивостью (выдерживают кипячение от нескольких минут до нескольких часов). Термоустойчивость обусловлена наличием специфического для эндоспор соединения - дипиколината кальция (отсутствует в вегетативной клетке, возникает непосредственно перед появлением термоустойчивости и исчезает при прорастании споры). Термоустойчивость также обуславливает малое содержание воды (на 20…35% меньше, чем в вегетативной клетке), наличие многочисленных оболочек, особенность кортекса, большое содержание липидов.
Эндоспоры присущи грамположительным бактериям (исключение составляют бактерии рода Desulfotomaculum). Как правило, извитые и кокковидные формы эндоспор не образуют (исключение род Sporosarcina).
Для бактерий спорообразование не является обязательным этапом жизненного цикла, так как при благоприятных условиях они могут длительное время развиваться без образования спор. Есть мутанты, не образующие спор.
Прорастание спор. Процесс прорастания протекает в несколько стадий (рисунок 1.7)
1. Активация прорастания отражает готовность спор к прорастанию. Она происходит в процессе старения споры (необратима), под воздействием редуцирующих веществ, повышенных температур, при снижении рН.
Основные свойства споры сохраняются, но увеличивается численность спор, способных к прорастанию. Наиболее часто используется тепловая обработка – прогрев спор в течение некоторого времени при высокой сублетальной температуре.
2. Инициация прорастания (необратима), при которой снижается устойчивость спор к прогреванию, краскам, высушиванию. Споры теряют термоустойчивость и светопреломление. Эти процессы сопровождаются выделением (до 30%) веществ споры – дипиколината кальция, глюкозамина, диаминопимелиновой кислоты, некоторых аминокислот. Низкомолекулярные белки споры разрушаются специальной споровой протеазой. Стадия длится несколько минут.
3. Собственно прорастание. В этой стадии происходит активный синтез белка и РНК, репликация ДНК, лизис оболочек споры, их разрыв в какой-нибудь точке споры и появление "ростка" новой клетки. Чаще всего спора прорастает полярно или латерально. Далее следует удлинение ростка и формирование полноценной вегетативной клетки. Процесс прорастания в среднем происходит за 2…3 ч.
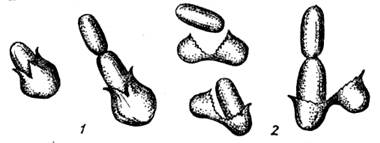
Рисунок 1.7 – Схема прорастания эндоспоры
1 - полярное прорастание; 2 – латеральное прорастание.
Цисты(лат. cista – ящик) - это округлые светопреломляющие образования, содержащие цитоплазму с нуклеоидом, окруженную ЦПМ с двумя оболочками - внутренней толстой и внешней многослойной. В цисту превращается вся бактериальная клетка. Цисты возникают в старых культурах. Они содержат много липидов. Цисты более устойчивы, чем вегетативные клетки к высушиванию, механическим воздействиям, лизоциму, слабо устойчивы к температуре. В благоприятных условиях (при наличии источника углерода) цисты прорастают. Цисты образуют некоторые метилотрофные бактерии, спирохеты, бактерии родов Azotobacter.
Движение бактерий
По способности передвигаться все бактерии делятся на две группы – неподвижные и подвижные (примерно 1/20 часть). Неподвижным бактериям размером менее 4 мкм присуще броуновское движение.
Движение поступательное с помощью жгутиков. Это наиболее распространенный тип движения.
 9
9
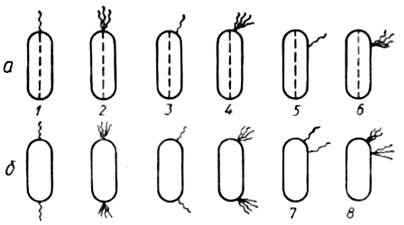
Рисунок 1.8 – Жгутики бактерий
a – жгутики расположены на одном конце клетки; б - жгутики расположены на обоих концах клетки; 1 – полюсные монотрихи; 2 – полюсные лофотрихи; б1 и б2 - амфитрихи 3 – подполюсные монотрихи; 4 – подполюсные лофотрихи; 5 – боковые монотрихи; 6 – боковые лофотрихи; 9 – перитрих.
По количеству и характеру расположения различают жгутики (рисунок 1.8):
-полюсные жгутики, когда один, два и более жгутиков расположены на одном или обоих полюсах клетки и основание жгутика обычно параллельно длинной оси клетки;
- подполюсные жгутики (или субполярные), когда один, два и более жгутиков расположены в месте перехода боковой поверхности в полюс клетки на одном или двух концах ее, и основание жгутика обычно составляет прямой угол с длинной осью клетки;
- боковые (или латеральные) жгутики, когда один, два и более жгутиков в виде кустика, расположены в средней точке одной из половин клетки;
- перитрихиальные жгутики, которые расположены по всей поверхности клетки по одному или пучками (обычно полюса клеток лишены их);
- смешанныежгутики, когда два или несколько жгутиков расположены в различных точках клетки.
Бактерии, имеющие один жгутик, называются монотрихами, пучок жгутиков – лофотрихами (или политрихами). Биполярные монотрихи или лофотрихи называются амфитрихами.
Количество жгутиков зависит от вида микроорганизма и условий культивирования. Жгутики очень тонкие образования, состоящие в основном (95%) из белка флагеллина (лат. flagellum– жгутик). Средняя длина жгутиков большинства бактерий 10…20 мкм. Лни могут составлять до 2% массы бактерий.
Большинство бактерий за секунду проходит расстояние, близкое длине их тела, но есть бактерии, движущиеся с большой скоростью (до 30 мкм/с). Скорость движения не зависит от количества жгутиков, но зависит от характера расположения жгутиков (бактерии с терминальным расположением жгутиков движутся быстрее, чем с перитрихиальным), зависит также от свойств среды (вязкости, температуры, рН, осмотического давления и пр.). Направленность движения обуславливается характером расположения жгутиков. Моно- и лофотрихи движутся прямолинейно (либо жгутиками вперед, либо жгутиками позади), перитрихи – беспорядочно, кувыркаются. Жгутики не являются жизненно необходимыми приспособлениями, их потеря не сопровождается изменением жизнедеятельности клетки.
Скользящий тип движения менее распространен у бактерий. Он присущ миксобактериям, некоторым микоплазмам, цитофагам. Скорость такого движения низкая (2…11 мкм/с). Большинство скользящих бактерий выделяет слизь.
Вращательный тип движения присущ извитым формам, например, спирохетам. Они передвигаются за счет колебания собственного тела, очень подвижны.
Таксичес
Дата добавления: 2017-11-21; просмотров: 7207;
Поиск по сайту
Узнать еще
- IV. Права и обязанности личного состава службы
- V. Подготовка личного состава службы
- VI. Тормоза подвижного состава
- VII. Клей разных составов
- VIII. Сигналы, применяемые для обозначения поездов, локомотивов и другого железнодорожного подвижного состава
- А. КРОВЬ 43. Состав крови
- Автоматизация составления технического задания и технической концепции на проектирование (подбор) муфты
- Автоматизированная система управления тяговым подвижным составом (ДИСТПС)

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
