Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 {corpus trapezoideum), волокна которого относятся к слуховому пути. В области трапециевидного тела располагается ядро, также имеющее отношение к слуховому пути, — nucleus dorsalis corporis trapezoidei.
{corpus trapezoideum), волокна которого относятся к слуховому пути. В области трапециевидного тела располагается ядро, также имеющее отношение к слуховому пути, — nucleus dorsalis corporis trapezoidei.
Вентральная часть моста содержит продольные и поперечные волокна, между которыми разбросаны собственные ядра серого вещества (nuclei pontis). Продольные волокна принадлежат к пирамидным путям (корково-мостовые волокна; fibrae corticopontinae), которые связаны с собственными ядрами моста, откуда берут начало поперечные волокна, идущие к коре мозжечка (мосто-мозжечковый путь; tractus pontocerebellaris).
Вся эта система проводящих путей связывает через мост кору полушарий большого мозга с корой полушарий мозжечка.
В дорзальной части моста находится его ретикулярная формация (formacio reticularis pontis), являющаяся продолжением такой же формации продолговатого мозга [240]. Поверх ретикулярной формации располагается выстланное эпендимой дно ромбовидной ямки с лежащими под ним ядрами черепных нервов.
В дорзальной части продолжаются также проводящие пути продолговатого мозга, располагающиеся между средней линией и nucleus dorsalis corporis trapezoidei и входящие в состав медиальной петли (lemniscus medialis). В медиальной петле перекрещиваются восходящие пути продолговатого мозга (tractus bul-bothalamicus).
Как указывалось выше, мост содержит многочисленные ядра. Ядра моста посылают волокна в кору противоположной стороны мозжечка. При этом они передают в кору мозжечка сведения о характере импульсов, идущих по корти-ко-бульбарным и кортико-спиннальным путям к нижележащим двигательным нейронам. Дор-зальный отдел моста (pars dorsalis pontis) содержит стволовые ядра черепно-мозговых нервов (V, VI, VIIи частично VIIIпары), ядра ретикулярной формации, а также восходящие (сенсорные) и нисходящие (двигательные) системы волокон. Дорзальный отдел моста называют также покрышкой моста (tegmentum pontis).
В мосту проходит пучок волокон, исходящий из нейронов центрального серого вещества среднего мозга (substantia grisea cenralis), a также от клеток zona incerta субталамической области, базальных ядер конечного мозга и красного ядра. Этот пучок связан с мозжечком посредством окончаний в нижней оливе, из которой полученная информация передается затем в кору мозжечка.
Получив по новым и старым двигательным путям информацию о положении и тонусе мышц, а также о предполагаемом двигательном акте, мозжечок перерабатывает ее и координирует двигательный акт. Импульсы направляются из мозжечка по эфферентным путям верхней
ножки. Такая координация может достигаться благодаря наличию связей мозжечка с ядрами ретикулярной формации среднего мозга или со зрительным бугром. По ним мозжечковые влияния проводятся к тем отделам коры мозга, где первоначально создается программа двигательного акта.
Мозжечок(cerebellum) (см. рис. 4.1.28). Мозжечок является производным заднего мозга, развившегося в связи с рецепторами гравитации. Поэтому он имеет прямое отношение к координации движений и является органом приспособления организма к преодолению основных свойств массы тела — тяжести и инерции. Развитие мозжечка в процессе филогенеза прошло три основных этапа соответственно изменению способов передвижения животного.
Мозжечок впервые появляется в классе круглоротых, у миног, в виде поперечной пластинки. У низших позвоночных (рыбы) выделяются парные ушковидные части (archicerebel-lum) и непарное тело (paleocerebellum), соответствующее червю. У пресмыкающихся и птиц сильно развито тело, а ушковидные части превращаются в рудиментарные. Полушария мозжечка возникают только у млекопитающих (neocerebellum), а у человека они достигают наибольшего развития.
Мозжечок помещается под затылочными долями полушарий большого мозга, дорзально от моста и продолговатого мозга. Лежит в задней черепной ямке. В нем различают объемистые боковые части, или полушария (hemispheria cerebellum), и расположенную между ними среднюю узкую часть — червь (vermis).
На переднем крае мозжечка находится передняя вырезка, которая охватывает прилежащую часть ствола мозга. На заднем крае имеется более узкая задняя вырезка, отделяющая полушария друг от друга [11, 397].
Поверхность мозжечка покрыта слоем серого вещества, составляющего кору мозжечка, и образует узкие извилины — листки мозжечка (folia cerebelli), отделенные друг от друга бороздами (fissurae cerebelli). Среди них самая глубокая fissura horizontalis cerebelli проходит по заднему краю мозжечка, отделяет верхнюю поверхность полушарий от нижней поверхности. С помощью горизонтальной и других крупных борозд вся поверхность мозжечка делится на ряд долек (lobuli cerebelli). Среди них необходимо выделить наиболее изолированную маленькую дольку — клочок (flocculus), лежащую на нижней поверхности каждого полушария у средней мозжечковой ножки, а также связанную с клочком часть червя — nodulus, узелок. Клочек соединен с узелком посредством тонкой полоски — ножки клочка (pedunculus flocculi), которая медиально переходит в тонкую полулунную пластинку — нижний мозговой парус (velum medullare inferius) (рис. 4.1.29).
Анатомия головного мозга

|
|
| 12 |
| 13 |
| 14 |
|
Рис. 4.1.29. Кора и ядра мозжечка:
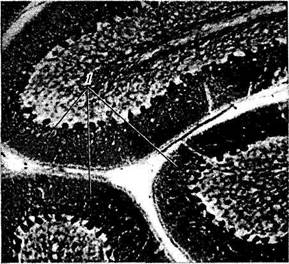
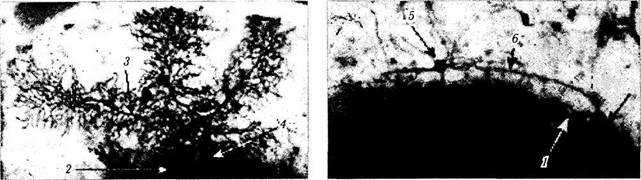
а — поперечный срез мозжечка (вид сверху) (/ — кора червя мозжечка; 2— пробковидное ядро; 3— шаровидное ядро; 4— ворота зубчатого ядра; 5 — ямочка мозжечка; 6 — верхний мозжечковый парус; 7 — верхние бугорки четверохолмия; 8 — нижние бугорки четверохолмия; 9 — верхние ножки мозжечка; 10 — ядро шатра; // — зубчатое ядро; 12 — ямочка зубчатого ядра; 13 — белое вещество; 14 — кора полушарий мозжечка; 15 — белое вещество); б, в, г—клеточная организация коры мозжечка (гистологические срезы коры полушарий мозжечка; импрегнация серебром) (/ — слой клеток Пуркине; 2 — слой гранулярных клеток; 3 — дендриты клеток Пуркине; 4 — клеточное тело; 5 — корзинкоподобная клетка; 6 — аксон)
В толще мозжечка располагаются парные ядра серого вещества, заложенные в каждой половине мозжечка среди белого ее вещества. По бокам от средней линии в области, где в мозжечок вдается шатер {fastigium), лежит самое медиальное ядро — ядро шатра (nucleus fastigii). Латеральнее от него расположено шаровидное ядро (nucleus globossus), а еще латеральнее — пробковидное ядро (nucleus embo-liformis). Наконец, в центре полушария находится зубчатое ядро (nucleus dentatus). Это ядро имеет вид серой извилистой пластинки, похожей на ядро оливы. Сходство зубчатого ядра мозжечка с имеющим также зубчатую форму ядром оливы не случайно, так как оба ядра связаны проводящими путями (оливо-моз-жечковый путь; tractus olivocerebellaris), и каждая извилина одного ядра аналогична извилине другого. Таким образом, оба ядра вмес-
те участвуют в осуществлении функции равновесия.
Названные ядра мозжечка имеют различный филогенетический возраст. Ядро шатра (nucleus fastigii) относится к самой древней части мозжечка — flocculus (archicerebellum), связанной с вестибулярным аппаратом. Пробковидное и шаровидное ядра (nucleus emboliformis et glo-bosus) относятся к старой части (paleo-cerebellum), возникшей в связи с движениями туловища. Зубчатое ядро (nucleus dentatus) относится к самой молодой (neocerebellum), развившейся в связи с передвижением при помощи конечностей. Поэтому при поражении каждой из этих частей нарушаются различные стороны двигательной функции, соответствующие различным стадиям филогенеза. Например, при повреждении флоккулонодулярной системы и ее ядра шатра нарушается равновесие тела
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
[325]. При поражении червя и соответствующих ему пробковидного и шаровидного ядер нарушается работа мускулатуры шеи и туловища, при поражении полушарий и зубчатого ядра — работа мускулатуры конечностей.
Белое вещество мозжечка на разрезе имеет вид мелких листочков растения, соответствующих каждой извилине, покрытых с периферии корой серого вещества. В результате общая картина белого и серого вещества на разрезе мозжечка напоминает дерево (arbor vitae cere-belli). Белое вещество мозжечка слагается из различного рода нервных волокон. Одни из них связывают извилины и дольки, другие идут от коры к внутренним ядрам мозжечка и, наконец, третьи связывают мозжечок с соседними отделами мозга. Эти последние волокна идут в составе трех пар мозжечковых ножек [4, б, 8, 9, 11] (рис. 4.1.30):
| 16 |
|
1. Нижние ножки (pedunculi cerebellares inferiores) (к продолговатому мозгу). В их составе задний спинно-мозжечковый путь (Флек-сига) (tractus spinocerebellaris posterior) идет к мозжечку от ядер задних канатиков продолговатого мозга, а также оливо-мозжечковый путь (tractus olivocerebellares). Оканчиваются эти
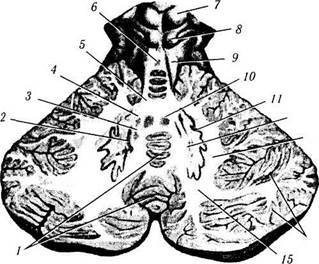
Рис. 4.1.30. Четвертый желудочек и мозжечок:
/ — задвижка; 2 — треугольник блуждающего нерва; 3 — треугольник подъязычного нерва; 4— тройничный бугорок; 5 — нижняя ямка; 6 — пограничная борозда; 7 — верхняя ямка; 8— латеральный карман; 9 — мозжечковые ножки (верхняя, средняя, нижняя); 10 — верхний мозговой парус; // — блоковый нерв; 12 — нижний бугорок; 13 — верхний бугорок; 14 — шишковидное тело; 15 — подушка зрительного бугра; 16 — третий желудочек; 17 — треугольник повока; 18 — коленчатые тела; 19 — задняя срединная борозда; 20 — верхняя мозжечковая ножка; 21 — голубоватое место; 22 — медиальное возвышение; 23 — колено лицевого нерва; 24 — вестибулярное поле; 25 — зубчатое ядро мозжечка; 26 — мозговые полоски; 27 — лента четвертого желудочка; 28 — бугорок клиновидного ядра; 29 —• бугорок тонкого ядра; 30 — задняя срединная борозда; 31—латеральный канатик; 32 — клиновидный пучок; 33 — тонкий пучок
пути в коре червя и коре полушарий. Кроме того, здесь идут волокна от ядер вестибулярного нерва, заканчивающиеся в ядре шатра. Благодаря всем этим волокнам мозжечок получает импульсы от вестибулярного аппарата и про-приоцептивного поля, вследствие чего становится ядром проприоцептивной чувствительности, совершающим автоматическую поправку на двигательную деятельность остальных отделов мозга. В составе нижних ножек идут также нисходящие пути в обратном направлении, а именно: от nucleus fastigii к латеральному вестибулярному ядру (nucleus vestibularis latera-lis), а от него — к передним рогам спинного мозга (преддверно-спинномозговой путь; tractus vestibulospinalis). При посредстве этого пути мозжечок оказывает влияние на спинной мозг.
2. Средние ножки (pedunculi cerebellares
medii) (к мосту). В их составе идут нервные
волокна от ядер моста к коре мозжечка. Воз
никающие в ядрах моста проводящие пути к
коре мозжечка (tractus pontocerebellaris) нахо
дятся на продолжении корково-мостовых путей
(fibrae corticopontinae), оканчивающихся в яд
рах моста после перекреста. Эти пути связыва
ют кору большого мозга с корой мозжечка, чем
и объясняется тот факт, что чем более развита
кора большого мозга, тем более развиты мост
и полушария мозжечка, что наблюдается у че
ловека.
3. Верхние ножки (pedunculi cerebellares su-
periores) (к крыше среднего мозга). Они состо
ят из нервных волокон, идущих в обоих направ
лениях: а) к мозжечку (передний спинно-моз
жечковый путь Говерса; tractus spinocerebel-
lares anterior) и б) от зубчатого ядра (nucleus
dentatus) мозжечка к покрышке среднего мозга
(мозжечково-покрышечный путь; tractus cere-
bellotegmentalis), который после перекреста
заканчивается в красном ядре и в таламусе. По
первым путям в мозжечок идут импульсы от
спинного мозга, а по вторым он посылает им
пульсы в экстрапирамидную систему, посред
ством которой сам влияет на спинной мозг.
Сложной структурой обладает кора мозжечка. Более подробно строение и функция мозжечка будут изложены, одновременно с вестибулярной системой в разделе, посвященном вестибулярному и оптокинетическому рефлексам.
Продолговатый мозг
Продолговатый мозг, или миеленцефа-лон (myelencephalon; medulla oblongata) (рис. 4.1.24; 4.1.31, см. цв. вкл.), представляет собой непосредственное продолжение спинного мозга в ствол головного мозга и является частью ромбовидного мозга. Он сочетает в себе черты строения спинного мозга и начального отдела головного, чем и оправдывается его название myelencephalon.
Анатомия головного мозга

 Продолговатый мозг имеет вид луковицы (bulbus cerebri — отсюда термин «бульбарные расстройства»). Его верхний расширенный конец граничит с мостом, а нижней границей служит место выхода корешков I пары шейных нервов или уровень большого отверстия затылочной кости [4, 6, 8, 11, 397, 578].
Продолговатый мозг имеет вид луковицы (bulbus cerebri — отсюда термин «бульбарные расстройства»). Его верхний расширенный конец граничит с мостом, а нижней границей служит место выхода корешков I пары шейных нервов или уровень большого отверстия затылочной кости [4, 6, 8, 11, 397, 578].
1. На передней (вентральной) поверхности
продолговатого мозга по средней линии про
ходит передняя срединная щель (fissura те-
diana anterior), являющаяся продолжением
одноименной борозды спинного мозга. По бо
кам ее на той и другой стороне находятся
два продольных тяжа — пирамиды (pyramides
medullae oblongatae), которые как бы про
должаются в передние канатики спинного моз
га. Составляющие пирамиды пучки нервных
волокон частично перекрещиваются в глуби
не передней срединной щели с аналогичными
волокнами противоположной стороны (decus-
satio pyramidum). После этого они спуска
ются в боковом канатике на другой стороне
спинного мозга, образуя латеральный корко-
во-спинномозговой путь (tractus corticospinalis
(pyramidalis) lateralis). Часть волокон оста
ются неперекрещенными и спускаются в пе
реднем канатике спинного мозга на своей сто
роне, формируя передний спинномозговой
путь (tractus corticospinalis (pyramidalis)
anterior).
Пирамиды отсутствуют у низших позвоночных и появляются по мере развития новой коры; поэтому они наиболее развиты у человека, так как пирамидные волокна соединяют кору большого мозга, достигшую у человека наивысшего развития, с ядрами черепных нервов и передними рогами спинного мозга.
Латерально от пирамиды лежит овальное возвышение — олива (oliva), которая отделена от пирамиды бороздкой (sulcus anterolateralis).
2. На задней (дорзальной) поверхности про
долговатого мозга тянется задняя срединная
щель (sulcus medianus posterior), являющаяся
непосредственным продолжением одноименной
борозды спинного мозга. По бокам щели лежат
задние канатики, ограниченные латерально с
той и другой стороны слабо выраженной задне-
латеральной щелью (sulcus posterolateralis).
По направлению кверху задние канатики рас
ходятся в стороны и идут к мозжечку, входя в
состав его нижних ножек (pedunculi cerebella-
res inferiores), окаймляющих снизу ромбовид
ную ямку.
Каждый задний канатик подразделяется при помощи промежуточной борозды на медиально расположенный тонкий пучок (fasciculus gra-cilis) и латеральный клиновидный пучок (fasciculus cuneatus). У нижнего угла ромбовидной ямки тонкий и клиновидный пучки приобретают утолщения (tuberculum gracilis и tuberculum cuneatum). Эти утолщения обусловлены наличием соименных с пучками ядрами серого ве-
щества. Это тонкое ядро (nucleus gracilis) и клиновидное ядро (nucleus cuneatus). В названных ядрах оканчиваются проходящие в задних канатиках восходящие волокна спинного мозга (тонкий и клиновидный пучки). Латеральная поверхность продолговатого мозга, находящаяся между переднелатеральной и заднелате-ральной бороздами (sulci posterolateralis et anterolateralis), соответствует боковому канатику. Из заднелатеральной борозды позади оливы выходят XI, X и IX пары черепных нервов. В состав продолговатого мозга входит нижняя часть ромбовидной ямки.
Продолговатый мозг возник в связи с развитием органов гравитации и слуха, а также в связи с жаберным аппаратом, имеющим отношение к дыханию и кровообращению. Поэтому в нем заложены ядра серого вещества, имеющие отношение к равновесию, координации движений, а также к регуляции обмена веществ, дыхания и кровообращения [4, 6, 8, 9, 11].
1. Ядро оливы (nucleus olivaris) имеет вид
извитой пластинки серого вещества, открытой
медиально (hilus), и обусловливает снаружи
выпячивание оливы. Оно связано с зубчатым
ядром мозжечка и является промежуточным
ядром равновесия.
2. Ретикулярная формация (fomatio reticu-
laris), образующаяся из переплетения нерв
ных волокон и лежащих между ними нервных
клеток.
3. Ядра VIII (частично), IX, X, XI (частично)
и XII пар черепно-мозговых нервов. Эти ядра
служат центрами регуляции дыхания и крово
обращения, центрами рвотного рефлекса и рав
новесия. Поэтому при повреждении продолго
ватого мозга может наступить смерть. Послед
няя функция реализуется вестибулярной вет
вью VIII пары черепно-мозговых нервов.
В дорзолатеральной части продолговатого мозга проходит в нисходящем направлении длинный пучок волокон тройничного нерва (V пара), который, по некоторым данным, достигает уровня 4-го шейного сегмента спинного мозга.
Белое вещество продолговатого мозга содержит длинные и короткие волокна. К длинным относятся проходящие транзитно в передние канатики спинного мозга нисходящие пирамидные пути, частично перекрещивающиеся в области пирамид. Кроме того, в ядрах задних канатиков (nuclei gracilis et cuneatus) находятся тела вторых нейронов восходящих чувствительных путей. Их отростки идут от продолговатого мозга к зрительному бугру (tractus bulbothalamicus). Волокна этого пучка образуют медиальную петлю (lemniscus medialis), которая в продолговатом мозге совершает перекрест (decussatio lemniscorum) и в виде пучка волокон, расположенных дорзальнее пирамид, между оливами — межоливный петлевой слой — идет далее.
Глава 4. ГОЛОВНОМ МОЗГ И ГЛАЗ
 Таким образом, в продолговатом мозге имеется два перекрестка длинных проводящих путей: вентральньный двигательный (decussatio pyramidum) и дорзальный чувствительный (decussatio lemniscorum).
Таким образом, в продолговатом мозге имеется два перекрестка длинных проводящих путей: вентральньный двигательный (decussatio pyramidum) и дорзальный чувствительный (decussatio lemniscorum).
К коротким путям относятся пучки нервных волокон, соединяющие между собой отдельные ядра серого вещества, а также ядра продолговатого мозга с соседними отделами головного мозга. Среди них следует отметить оливо-моз-жечковый путь (tractus olivocerebellaris) и лежащий дорзально от межоливного слоя медиальный продольный пучок (fasciculum longi-tudinalis medialis) (см. рис. 4.1.26).
Топографические взаимоотношения главнейших образований продолговатого мозга видны на поперечном срезе, проведенном на уровне олив. Отходящие от ядер подъязычного и блуждающего нервов корешки делят продолговатый мозг на той и другой стороне на три области: заднюю, боковую и переднюю. В задней лежат ядра заднего канатика и нижние ножки мозжечка, в боковой — ядро оливы и ретикулярная формация и в передней — пирамиды.
Через продолговатый мозг проходят пути болевой, температурной и тактильной чувствительности. Собраны эти волокна в спиральную петлю (lemniscus spiralis) и медиальную петлю (lemniscus medialis) (рис. 4.1.32, см. цв. вкл.). Волокна медиальной петли начинаются от клеток, расположенных в дорзальных отделах кау-дальной части продолговатого мозга. Система волокон медиальной петли связана с дискриминационной чувствительностью, чувством положения и вибрации.
Нарушение функции продолговатого мозга приводит к тяжелым последствиям. Продолговатый мозг сдавливается в тех случаях, когда мозжечковые миндалины проникают через большое отверстие (foramen magnum) при повышении внутричерепного давления. Образование грыжи приводит к парезам с потерей чувствительности и нередко сопровождается вертикальным нистагмом.
Нарушение функции мозга встречается и в том случае, когда нижняя часть мозжечка проникает через большое отверстие при пороке развития Арнольда — Хиари. При сирингобуль-бии появляется полость в продолговатом мозге, вызывая нистагм и атрофию языка в результате нарушений функций ядер подъязычного нерва.
Ретикулярная формация.В предыдущих разделах неоднократно упоминалось о ретикулярной формации (formatio reticularis) (см. рис. 4.1.31).
Под этим названием подразумевают совокупность структур, расположенных в центральных отделах мозгового ствола [4, 6, 8, 9, 11].
Образована ретикулярная формация группами мелких, средних и крупных мультиполярных вставочных нейронов с различным характером
ветвления дендритов и аксонов. Используют эти нейроны различные нейромедиаторы и окружены сетями нервных волокон. Ретикулярная формация простирается от продолговатого до промежуточного мозга, достигая в краниальном направлении своего наибольшего развития [240]. При этом диффузное распределение нейронов сменяется более компактным их расположением с формированием отдельных ядер, часть которых приведена на рис. 4.1.31.
Ретикулярная формация характеризуется следующими морфологическими особенностями:
1. Нейроны ретикулярной формации отлича
ются наличием коротких и слабо ветвящихся
дендритов. Аксоны, наоборот, длинные и делят
ся на восходящую и нисходящую ветви. Аксоны
отдают многочисленные коллатерали, благода
ря чему аксон может контактировать с огром
ным числом нервных клеток.
2. Нервные волокна идут в самых различ
ных направлениях, напоминая под микроскопом
сеть. Эта особенность послужила основанием
для Дейтерса назвать это скопление нейронов
сетчатой, или ретикулярной формацией.
3. Клетки ретикулярной формации местами
рассеяны, а местами образуют ядра. В настоя
щее время описано около 100 отдельных ядер.
В зависимости от типа используемых нейронами медиаторов в передаче нервного импульса различают три группы скопления нейронов. Нейромедиатором первой группы нейронов является дофамин. Расположены эти нейроны с вентральной стороны области покрышки, а их отростки направляются к лимбической системе и обонятельному тракту.
Скопления нейронов, использующих норэпи-нефрин, рассеяны на большом протяжении (locus coeruleus). Связаны они с корой большого и малого мозга, зрительным бугром, лимбической системой, стволом мозга и спинным мозгом [378].
Третья группа нейронов относится к серо-тонинэргическим. Формируют они ядро шва (п. raphe) в среднем мозге, мосту и медуллярной части ретикулярной формации. Эти нейроны связаны с лимбической системой, обонятельной областью и, в меньшей степени, мозжечком и спинным мозгом [240, 271,403, 404].
Область распространения ретикулярной формации точно еще не установлена. На основании физиологических данных, она расположена по всей длине мозгового ствола и занимает центральное положение в продолговатом мозге, мосте, среднем мозге, в гипоталамической области и даже в медиальной части таламуса. Как филогенетически более древняя ретикулярная формация локализуется в покрышке ствола головного мозга.
Ретикулярная формация связана со всеми отделами центральной нервной системы. Различают следующие связи:
Анатомия головного мозга
1.  Ретикулопетальные связи, идущие от мо
Ретикулопетальные связи, идущие от мо
торных и сенсорных областей коры мозга, зри
тельного бугра и гипоталамуса.
2. Ретикулофугальные связи, идущие к се
рому веществу и ядрам головного и спинного
мозга.
3. Ретикулоретикулярные связи (восходя
щие и нисходящие) между различными ядрами
самой ретикулярной формации.
Ретикулярная формация оказывает общее неспецифическое активизирующее воздействие на всю кору головного мозга, что обеспечивается наличием восходящих проводящих путей от сетчатой формации ко всем долям мозговых полушарий. Поэтому ее называют также восходящей активирующей ретикулярной системой. Будучи связанной коллатералями аксонов своих клеток со всеми проходящими через ствол мозга специфическими афферентными проводящими путями, она получает от них импульсы и несет неспецифическую информацию в мозговую кору.
В результате этого процесса через мозговой ствол проходят в кору мозга две афферентные системы. Первая система является специфической и несет она все специфические чувствительные проводящие пути, несущие импульсы от рецепторов, котрые заканчиваются на телах клеток преимущественно IV слоя коры. Вторая система неспецифическая. Заканчивается она на дендритах всех слоев коры. Взаимодействие обеих этих систем обусловливает окончательную реакцию корковых нейронов. Таково современное представление о двух афферентных системах головного мозга.
Различные функции ретикулярной формации изучены далеко не полностью. К наиболее важным из них относятся: 1. Регуляция уровня сознания путем воздействия на активность нейронов коры мозга (например, участие в цикле сон/бодрствование). 2. Придание аффективно-эмоциональной окраски сенсорным стимулам, в том числе болевым сигналам, путем проведения афферентной информации к лимбической системе. 3. Регуляция вегетативных функций (цир-куляторные, дыхательные, акт глотания и др.). 4. Участие в позных и целенаправленных движениях в качестве важного компонента двигательных центров ствола головного мозга, включая движения глаза [58, 572].
Спинной мозг
Спинной мозг (medulla spinalis) (рис. 4.1.33, 4.1.34) располагается в позвоночном канале и имеет вид округлого тяжа, расширенного в шейном и поясничном отделах. В центре его расположен канал. Спинной мозг состоит из двух симметричных половин, разделенных спереди передней срединной щелью (fissura me-diana anterior), а сзади — задней срединной бороздой (sulcus medianus posterior).
Спинной мозг характеризуется сегментарным строением. С каждым сегментом связана пара передних (вентральных) и пара задних (дорзальных) корешков [4, 6, 8, 9, 11, 397].
В спинном мозге сосредоточены нервные клетки, аксоны которых дают начало нервам, идущим к поперечнополосатым мышцам тела. Эти, а также другие нейроны образуют клеточную «сердцевину» спинного мозга (серое вещество), расположенную вокруг его центрального канала. На поперечных срезах серое вещество спинного мозга имеет форму буквы Н. В нем различают парные передние (вентральные) (cornu anterior) и задние (дорзальные; соти posterior) рога, а также промежуточную соединяющую их часть (центральное промежуточное серое вещество; substancia (grisea) intermedia centralis) (рис. 4.1.33).
Задние рога выполняют главным образом сенсорные функции и содержат нейроны, которые передают чувствительные импульсы в лежащие выше центры или к расположенным вентральнее двигательным клеткам, замыкая таким образом рефлекторную дугу [3].
В передних рогах находятся нейроны, ин-нервирующие мышцы. В промежуточной зоне спинного мозга находятся клетки, связывающие его чувствительный и двигательный отделы.
С 1-го грудного сегмента спинного мозга и примерно до третьего поясничного сегмента в боковых отделах серого вещества расположена клеточная зона, занимающая промежуточное положение между передними и задними рогами. Это так называемый боковой столб (латеральные рога; columna lateralis). В его состав входит так называемый боковой промежуточный столб (columna intermedia lateralis [auto-nimica]), определяемый на протяжении всех грудных и первых двух поясничных сегментов спинного мозга (Tl—L2). Содержит он преганг-лионарные нейроны вегетативной нервной системы.
Нейроны располагаются в сером веществе в виде скоплений (ядер), в которых происходит переключение импульсов с клетки на клетку. На основании расположения нейронов, их цитологических особенностей, характера связей и функции выделено десять пластин, идущих в ростро-каудальном направлении. В зависимости от топографии аксонов нейроны спинного мозга подразделяют на следующие типы: 1. Корешковые нейроны, аксоны которых образуют передние корешки. 2. Внутренние нейроны, отростки которых заканчиваются в пределах серого вещества спинного мозга. 3. Пучковые нейроны, аксоны которых образуют пучки волокон в белом веществе спинного мозга в составе приводящих путей.
Задние рога содержат несколько ядер, образованных мультиполярными вставочными нейронами, на которых оканчиваются аксоны псев-
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
доуниполярных нейронов спинальных ганглиев, а также волокна нисходящих путей из лежащих выше центров. В задних рогах выявляется высокая концентрация таких нейромедиаторов, как серотонин, энкефалин, вещество Р.
Аксоны вставочных нейронов оканчиваются в сером веществе спинного мозга на мотонейронах, лежащих в передних рогах. Они также образуют межсегментальные связи в пределах серого вещества спинного мозга, а также выходят в белое вещество спинного мозга, где
образуют восходящие и нисходящие проводящие пути. Часть аксонов при этом переходит на противоположную сторону спинного мозга [11, 397].
Боковые рога хорошо выражены на уровне грудных и крестцовых сегментов спинного мозга и содержат ядра, образованные телами вставочных нейронов, которые относятся к симпатическому и парасимпатическому отделам вегетативной нервной системы. На денд-ритах и телах этих клеток оканчиваются аксо-
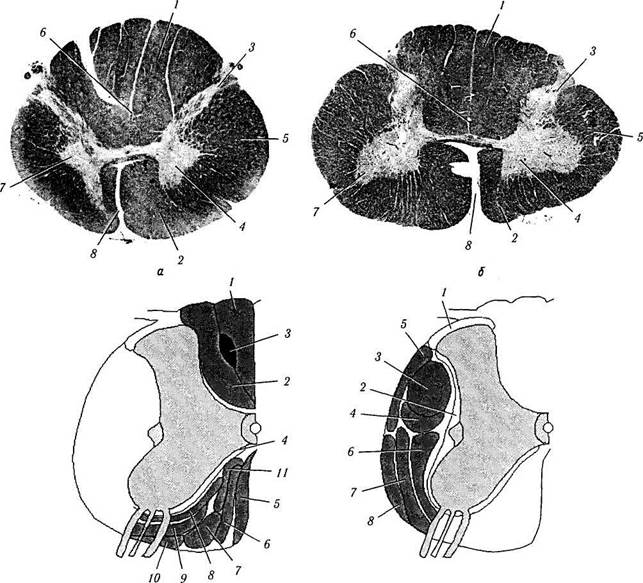
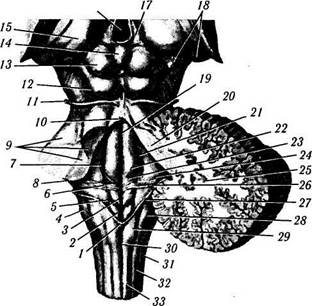
Рис. 4.1.33. Строение спинного мозга:
а, б — гистологический срез спинного мозга на уровне СЗ (а) и С8 (б) (1—дорзальный канатик; 2— вентральный канатик; 3— дорзальный рог; 4— вентральный рог; 5 — латеральный канатик; 6 — центральный канал; 7— промежуточное серое вещество; 8— передняя срединная щель); в — проводящие пути переднего и заднего канатиков (/ — нежный пучок; 2— клиновидный пучок; 3— тракт Шультце; 4— собственный пучок; 5 — передний (прямой) кортикоспинальный тракт; 6 — тектоспинальный тракт; 7 — медиальный продольный пучок; 8— ретикулоспинальныи тракт; 9 — спинноталамический тракт; 10 — оливоспинальный тракт; // — вестибулоспинальный тракт); г — боковой канатик (/—дорзолатеральный пучок; 2 — собственный пучок; 3 — латеральный (перекрещенный) кортикоспинальный тракт; 4 — руброспинальный тракт; 5 — задний спинно-мозжечковый тракт; 6—передний спинно-мозжечковый тракт; 7 — спинноталамический тракт; 8 — ретикулоспинальныи тракт)
Анатомия головного мозга
 |  |
|
|
| 10 1 |
| 14 |
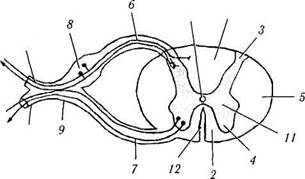
Рис. 4.1.34. Схематическое изображение спинного мозга и рефлекторных дуг:
а — спинной мозг и периферический нерв (/ —дорзальный канатик; 2 — вентральный канатик; 3 — дорзальный рог; 4 — вентральный рог; 5 — латеральный канатик; 6 — дорзальный корешок; 7 — вентральный корешок; 5 — дорзальный корешок ганглия; 9 — спинной нерв; 10 — центральный канал; // — промежуточное серое вещество; 12 — передняя срединная щель; 13 — дорзальная веточка; 14 — вентральная веточка); б — схема моносинаптической рефлекторной дуги (/ — рецептор; 2— эфферентное волокно; 3 — рефлекторный центр; 4 — афферентное волокно; 5 — эффекторный орган); в — схема формирования симпатической рефлекторной дуги (/—латеральный рог; 2 — спинной нерв; 3 — паравертебральный ганглий; 4 — превертебраль-ный ганглий; 5 — белые соединительные веточки; 6 — серые соединительные веточки)
ны а) псевдоуниполярных нейронов, несущих импульсы от рецепторов, расположенных во внутренних органах, б) нейронов центров регуляции вегетативных функций, тела которых располагаются в продолговатом мозге. Аксоны вегетативных нейронов выходят из спинного мозга в составе передних корешков, образуют преганглионарные волокна, направляющиеся к симпатическим и парасимпатическим узлам. В нейронах боковых рогов основным медиатором является ацетилхолин. Выявляется также ряд нейропептидов — энкефалин, нейротензин, вазоинтерстициальный кишечный полипептид (VIP), вещество Р, соматостатин, пептид, связанный с кальцитониновым геном. Передние рога содержат мультиполярные двигательные клетки (мотонейроны), объединенные в ядра. Различают крупные (диаметр тела 35—70 мкм) альфа-мотонейроны и рассеянные между ними
более мелкие (15—35 мкм) гамма-мотонейроны. На отростках и телах мотонейронов имеются многочисленные синапсы, оказывающие на них возбуждающие и тормозные воздействия. На мотонейронах оканчиваются: а) коллатера-ли аксонов псевдоуниполярных клеток спиналь-ных узлов, образующие с ними двухнейронные (моносинаптические) рефлекторные дуги; б) аксоны вставочных нейронов, тела которых лежат в задних рогах спинного мозга; в) аксоны клеток Реншоу, образующие тормозные аксо-соматические синапсы; г) волокна нисходящих путей пирамидной и экстрапирамидной систем, несущие импульсы из коры большого мозга и ядер ствола мозга. Гамма-мотонейроны, в отличие от альфа-мотонейронов, не имеют непосредственной связи с чувствительными нейронами спинальных узлов. Аксоны альфа-мотонейронов отдают коллатерали, оканчивающиеся на телах вставочных нейронов (клетки Реншоу), и покидают спинной мозг в составе передних корешков, направляясь в составе смешанных нервов к соматическим мышцам, на которых они оканчиваются нервно-мышечными синапсами. Более тонкие аксоны гамма-мотонейронов имеют такой же ход и образуют окончания на интра-фузальных волокнах нервно-мышечных веретен.
Серое вещество спинного мозга окружено слоем белого вещества (substantia alba), состоящего из восходящих и нисходящих миелинизи-рованных волокон.
Белое вещество разделяется серым веществом на симметричные дорзальные, латеральные и вентральные канатики (fasciculus anterior, lateralis et ventralis). Они состоят из продольно идущих нервных волокон, образующих восходящие и нисходящие пути. Для каждого тракта характерно преобладание волокон, образованных однотипными нейронами. По этой причине пути отличаются и нейромедиаторами. В соответствии с этим различают моноамин-эргические, холинэргические, ГАМК-эргичес-кие, глютаматэргические, глицинэргические и пептидэргические пути. Проводящие пути включают две группы: проприоспинальные и супра-спинальные пути.
Проприоспинальные пути являются собственно проводящими путями спинного мозга. Образованы они аксонами вставочных нейронов, которые осуществляют связь между его различными отделами. Эти пути проходят в основном на границе серого и белого веществ в составе латеральных и вентральных канатиков.
Супраспинальные проводящие пути обеспечивают связь спинного мозга со структурами головного мозга и включают восходящие спинноцеребральные и нисходящие цереброспинальные пути. Спинноцеребральные пути обеспечивают передачу в головной мозг сенсорной информации от спинного мозга, а цереброспинальные— от головного мозга спинному. К наи-
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 более важным путям относятся пирамидная и экстрапирамидная системы. Пирамидная система образована аксонами пирамидных клеток большого мозга, которые на уровне продолговатого мозга переходят на противоположную сторону и формируют латеральный и вентральный кортикоспинальные тракты. Пирамидная система контролирует точные произвольные движения скелетной мускулатуры, в особенности конечностей. Экстрапирамидная система (см. выше) образована нейронами ядер среднего, продолговатого мозга и моста. Их не
более важным путям относятся пирамидная и экстрапирамидная системы. Пирамидная система образована аксонами пирамидных клеток большого мозга, которые на уровне продолговатого мозга переходят на противоположную сторону и формируют латеральный и вентральный кортикоспинальные тракты. Пирамидная система контролирует точные произвольные движения скелетной мускулатуры, в особенности конечностей. Экстрапирамидная система (см. выше) образована нейронами ядер среднего, продолговатого мозга и моста. Их не
Дата добавления: 2016-06-05; просмотров: 2504;
Поиск по сайту
Узнать еще
- I. Синусы твердой оболочки головного мозга.
- II. Анатомия продолговатого мозга, ретикулярная формация
- IV. проводящие пути головного и спинного мозга
- V Патопсихологическое – при нарушении целостности мозга происходит нарушение психической деятельности
- VII. Вены глазницы.
- А Нейропсихологические синдромы поражения задних отделов коры больших полушарий головного мозга.
- Абсцессы головного мозга. Хирургическое лечение
- Альтернирующие синдромы при поражении ствола головного мозга

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
