ИММУННАЯ СИСТЕМА ЧЕЛОВЕКА
Иммунная система включает специализированную, анатомически обособленную лимфоидную ткань, «разбросанную» по всему организму в виде различных лимфоидных образований и отдельных клеток.
Различают первичные - центральные (костный мозг и тимус) и вторичные - периферические (селезенка, лимфатические узлы, скопления лимфоидной ткани) органы иммунной системы. Все они взаимосвязаны системой кровообращения, лимфотока и единой системой иммунорегуляции.
Первичные - центральные органы иммунной системы
Центральные органы иммунной системы - костный мозг и тимус выполняют важнейшие функции, обеспечивая самообновление иммунной системы, в этих органах идут процессы пролиферации клеток предшественников, их дифференцировка и созревание, вплоть до выхода в циркуляцию и заселения периферических органов иммунной системы зрелыми иммунокомпетентными клетками.
Рис. 4. Клетки крови формируются в костном мозге (фотография).
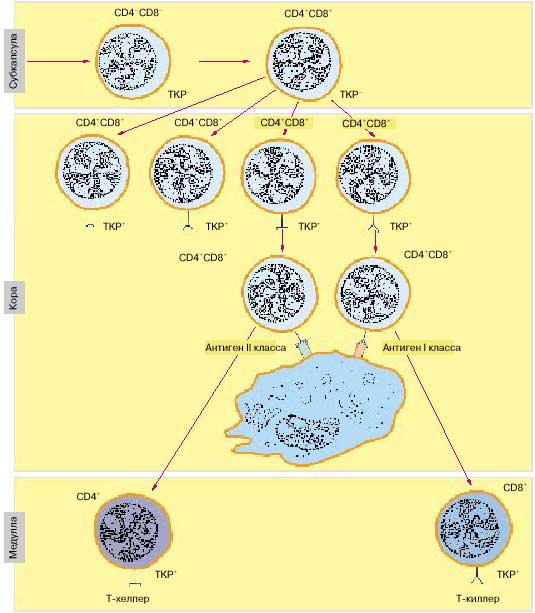
Рис. 5. В тимусе происходит созревание Т-хелперов и Т-киллеров, которые изначально поступают из костного мозга незрелыми, в процессе созревания они приобретают ТКР для распознавания антигена и CD4+ или CD8+ рецепторы для согласованной работы с другими иммуноцитами (схема).
Вторичные - периферические органы иммунной системы
Периферические органы иммунной системы - лимфатические узлы, селезенка и лимфоидная ткань, ассоциированная со слизистыми - являются местом встречи антигенов с иммунокомпетентными клетками, местом распознавания антигена и развития специфического ответа, местом взаимодействия иммунокомпетентных клеток, их пролиферации (клональной экспансии), антиген-зависимой дифференцировки и местом накопления продуктов иммунного ответа.
АНТИТЕЛА
Антитела (иммуноглобулины) (АТ, Ig) - в большинстве случаев представлены сывороточными гликопротеинами, входящие в состав фракции гамма-глобулинов. АТ образуют один из основных классов белков крови, составляя 20% массы белка плазмы. Молекула АТ состоит из 2 идентичных тяжелых (Н-цепи) и 2 идентичных легких (L-цепи) аминокислотных цепей, соединенных дисульфидными -S-S- связями (рис.3). В цепях различают вариабельную область (V-область) в N-концевой части и постоянную, или константную, область (С-область). V-область у разных АТ варьирует. V-области L- и Н-цепей образуют Аг-связывающий центр(активный центр АТ, паратоп), или Fab-фрагмент. Константная область молекулы имеет Fc-фрагмент.Аг-связывающий центр, образованный вариабельными областями L- и Н-цепей; именно с ним взаимодействует Аг-детерминанта (эпитоп) антигена. Аг-связывающий центр комплементарен эпитопу Аг по приципу «ключ-замок». Молекула Ig как минимум двухвалентна; такие антитела известны как полные АТ,а мономеры с меньшей валентностью - как неполные АТ. Fc-фрагмент определяет специфичность связывания молекулы Ig с клетками-эффекторами (например, макрофагами, полиморфноядерными лейкоцитами, тучными клетками), несущими на своей поверхности рецепторы Fc-фрагмента.
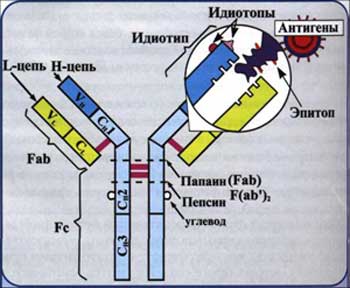
Рис. 6. Структура иммуноглобулина.
Классы иммуноглобулинов и свойства антител
В зависимости от структуры Н-цепей выделяют 5 разных классов (изотипов) АТ: IgA, IgD, IgE, IgG и IgM.
1. IgM -пентамер из 5 субъединиц, соединенных дисульфидными связями, имеет 10 Аг-связывающих участков. IgM - наиболее ранний класс АТ, обнаруживаемый при первичном попадании Аг в организм, т.е. наличие IgM к Аг конкретного возбудителя указывает на наличие острого инфекционного процесса. Молекулы IgM опсонизируют, агглютинируют, преципитируют и лизируют содержащие Аг структуры, а также легко активируют систему комплемента.
2. IgG - Основной класс АТ (до 75% всех Ig), защищающих организм от бактерий, вирусов и токсинов. После первичного контакта с Аг синтез IgM обычно сменяется образованием более дифференцированных IgG. Максимальные титры IgG при первичном ответе наблюдаются на 6-8 сутки. Обнаружение высоких титров IgG к Аг конкретного возбудителя указывает на то, что организм находится на стадии реконвалесценции или конкретное заболевание перенесено недавно.
IgG непосредственно участвуют в реакциях иммунного цитолиза, реакциях нейтрализации, а также усиливают фагоцитоз. Только IgG беременной форсируют плацентарный барьер, проходят черезбазальную мембранутрофобласта в соединительную ткань плода и попадает в капилляры плода. Транспорт IgG через плаценту обеспечивает формирование пассивного иммунитета у плода.
3. IgA (в сывороточном пуле Ig составляют 15-20%) секретируются на поверхность эпителиев, присутствуют в слюне, слезах, молоке, выделяются на поверхность слизистых оболочек, где взаимодействуют с Аг, усиливая защитные свойства слизистых оболочек пищеварительного тракта, дыхательных, половых и мочевыделительных путей. В сыворотке IgA циркулируют в виде мономеров, а в секретируемых SIgA преобладают четырехвалентные димеры, содержащие одну J-цепь и дополнительную полипептидную цепь (синтезируемых эпителиальными клетками секреторный компонент).
4. IgE специфически взаимодействуют с тучными клетками и базофильными лейкоцитами; эти клетки содержат многочисленные гранулы с биологически активными аминами. Защитные потенции IgE направлены преимущественно против гельминтов (нематод).
5. IgD. Биологическая роль этого Ig не установлена; IgD обнаруживают на поверхности развивающихся В-лимфоцитов; в сыворотке здоровых лиц он присутствует в крайне низкой концентрации.
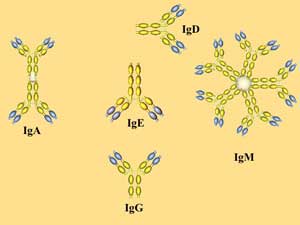
Рис. 7. Антитела разных классов отличаются биологическими свойствами.
Защитные механизмы антител (функции).
АТ помогают предотвратить инфицирование и/или элиминировать возбудитель, активируя при этом несколько механизмов.
- Опсонизация (иммунный фагоцитоз) - связывание АТ с клеточной стенкой микроорганизма с последующим эффективным поглощением образовавшегося комплекса фагоцитом при взаимодействии Fc-фрагмента АТ с соответствующим Fc-рецептором (FcR) на мембране фагоцита.
- Антитоксический эффект. АТ могут связывать и тем самым инактивировать бактериальные токсины.
- Активация комплемента. Антитела (IgM и IgG) после связывания с Аг (микроорганизм, опухолевая клетка и др.) могут активировать каскад сывороточных протеаз (систему комплемента), что приводит к уничтожению индуктора в результате перфорации его клеточной стенки, усилению хемотаксиса и иммунного фагоцитоза.
- Нейтрализация. Взаимодействуя с рецепторами клетки, связывающими бактерии или вирусы, АТ могут препятствовать адгезии и проникновению микроорганизмов в клетки организма-хозяина.
- Антителозависимаяцитотоксичность.Распознавание клетки-«мишени» антителами, адсорбированными на Fc-рецепторах NK-клеток, с последующим разрушением их.
Динамика антителообразования
Первичный ответ.Появлению АТ предшествует латентный период продолжительностью 3-5 суток. В это время происходит распознавание Аг и образование клонов плазматических клеток. Затем наступает логарифмическая фаза, соответствующая поступлению АТ в кровь; ее продолжительность - 7-15 суток. Постепенно титры АТ достигают пика и наступает стационарная фаза, продолжительнстью 15-30 суток. Ее сменяет фаза снижения титров АТ, длящаяся 1-6 месяцев.
Вторичный ответ. После антигенной стимуляции часть В- и Т-лимфоцитов циркулирует в виде клеток памяти. Особенности вторичного иммунного ответа - высокая скорость антителообразования, появление максимальных титров АТ и длительное (иногда многолетнее) их циркулирование. Основные характеристики вторичного ответа:
· образование АТ индуцируется значительно меньшими дозами Аг;
· индуктивная фаза сокращается до 5-6 ч;
· среди АТ доминируют IgG с большой аффинностью, пик их наступает раньше (3-5 сут);
· АТ образуются в более высоких титрах и циркулируют в организме длительное время.
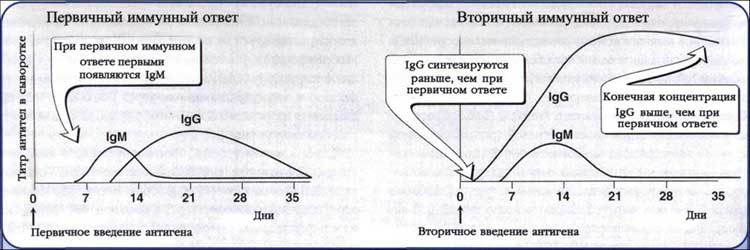
Рис. 8. Антителобразование при первичном и вторичном иммунном ответе.
ФОРМЫ ИММУНИТЕТА
Специфический иммунный ответ развивается в организме параллельно с развитием инфекции или после вакцинации и приводит к формированию ряда специфических эффекторных механизмов противоинфекционной защиты:
1. Гуморальный иммунный ответ (В-лимфоцит);
2. Клеточный иммунный ответ (Т-лимфоцит);
3. Иммунологическая память (Т- и В-лимфоциты);
4. Иммунологическая толерантность.
К этим механизмам относятся эффекторные молекулы (антитела) и эффекторные клетки (Т-лимфоциты и макрофаги) иммунной системы.
Гуморальные иммунные реакции
В гуморальных иммунных реакциях участвуют три клеточных типа: макрофаги (Аг-представляющие клетки), Т-хелперы и В-лимфоциты.
Аг-представляющие клетки фагоцитируют микроорганизм и перерабатывают его, расщепляя на фрагменты (процессинг Аг). Фрагменты Аг выставляются на поверхности Аг-представляющей клетки вместе с молекулой МНС. Комплекс «Аг-молекула МНС класса II» предъявляется Т-хелперу. Распознавание комплекса Т-хелпером стимулирует секрецию ИЛ-1 макрофагами.
Т-хелпер под действием ИЛ-1 синтезирует ИЛ-2 и рецепторы к ИЛ-2; последний стимулирует пролиферацию Т-хелперов, а также ЦТЛ. Таким образом, после взаимодействия с Аг-представляющей клеткой Т-хелпер приобретает способность отвечать на действие ИЛ-2 бурным размножением. Биологический смысл этого явления состоит в накоплении Т-хелперов, обеспечивающих образование в лимфоидных органах необходимого пула плазматических клеток, вырабатывающих АТ к данному Аг.
В-лимфоцит. Активация В-лимфоцита предполагает прямое взаимодействие Аг с молекулой Ig на поверхности В-клетки. В этом случае сам В-лимфоцит перерабатывает Аг и представляет его фрагмент в связи с молекулой МНС II на своей поверхности. Этот комплекс распознает Т-хелпер, отобранный при помощи того же Аг. Узнавание рецептором Т-хелпера комплекса Аг-молекула МНС класса II на поверхности В-лимфоцита приводит к секреции Т-хелпером ИЛ-2, ИЛ-4, ИЛ-5, ИЛ-6, под действием которых В-клетка размножается, образуя клон плазматических клеток (плазмоцитов). Плазмоциты синтезируют антитела. Часть зрелых В-лимфоцитов после антигензависимой дифференцировки циркулируют в организме в виде клеток памяти.
Антитела, специфически взаимодействуя с антигенными детерминантами (эпитопами) на поверхности микроорганизмов, образуют с ними иммунные комплексы, что ведет к активации мембраноатакующего комплекса системы комплемента и лизису микробных клеток. Кроме того, иммунные комплексы, включающие микроорганизмы и специфические антитела, быстрее и легче захватываются фагоцитирующими клетками организма при участии Fc-рецепторов. При этом ускоряется и облегчается внутриклеточная гибель и переваривание. Защитная роль антител в антитоксическом иммунитете определяется также их способностью нейтрализовать токсины. Секреторные иммуноглобулины класса А обеспечивают местный специфический иммунитет слизистых оболочек, препятствуя прикреплению и проникновению патогенных микроорганизмов.
Вместе с тем гуморальная защита малоэффективна против внутриклеточно паразитирующих бактерий, риккетсий, хламидий, микоплазм, грибов, простейших и вирусов. Против этих возбудителей более эффективны клеточные механизмы специфического иммунитета, к которым относится иммунное воспаление - реакция гиперчувствительности замедленного типа (ГЗТ) и цитотоксическая активность Т-киллеров, NК-клеток, макрофагов.
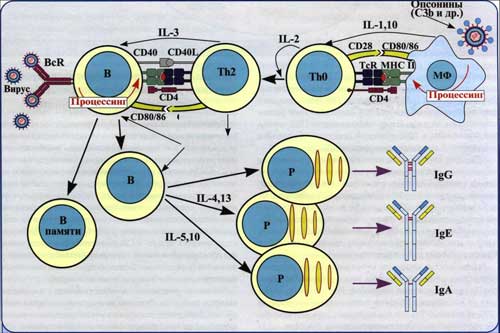
Рис. 1. Гуморальный иммунный ответ. В результате кооперации макрофагов, Т-хелперов и В-лимфоцитов и дальнейшей дифференцировки В-лимфоцитов в плазматические клетки, последние продуцируют антитела, которые нейтрализуют антиген.
Клеточные иммунные реакции
В очаге иммунного воспаления Т-эффекторы ГЗТ, активированные при контакте с микробными антигенами, продуцируют лимфокины, индуцирующие микробоцидные механизмы фагоцитов. В результате усиливается внутриклеточная гибель захваченных фагоцитами возбудителей.
Гибель клеток-«мишеней» вместе с паразитирующими в них возбудителями может наступить вследствие их распознавания Т-киллерами, специфически сенсибилизированных против микробных антигенов.
Другой механизм гибели зараженных клеток носит название антителозависимой цитотоксичности (АЗЦТ). Он заключается в распознавании микробных антигенов на мембране зараженной клетки-«мишени» антителами, адсорбированными на Fc-рецепторах NK-клеток или макрофагов. При этом цитотоксичность является результатом действия лизосомных ферментов и других продуктов секреции данных клеток.
В целом клеточные механизмы обеспечивают защиту организма против факультативно и облигатно внутриклеточных паразитов, что позволяет оценивать напряженность специфического иммунитета по результатам кожно-аллергической реакции. Этим же объясняется и тот факт, что наиболее эффективными для специфической профилактики таких инфекций являются вакцины из живых ослабленных микроорганизмов, активирующие клеточные механизмы иммунитета.
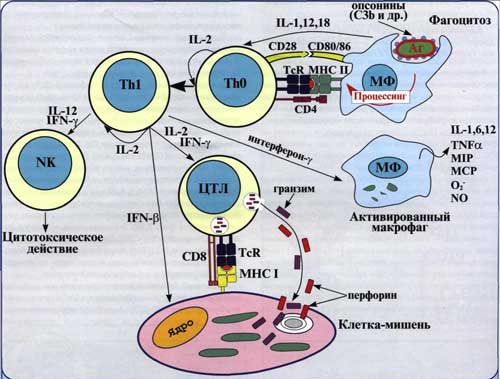
Рис. 2. Клеточный иммунный ответ опосредован активированными Т-хелперами макрофагами и другими фагоцитирующими клетками, а также цитотоксическими Т-лимфацитами.
Иммунологическая память
Иммунологическая память - способность организма отвечать на повторное введение антигена иммунной реакцией, характеризующейся большей силой и более быстрым развитием.
Клетки иммунологической памяти - долгоживущие Т- и В-лимфоциты, сохраняющие многие годы способность реагировать на повторное введение антигена, так как вырабатываются рецепторы к этому антигену. Иммунологическая память проявляется как ускоренный специфический ответ на повторное введение антигена.
Иммунологическая память к антигенным компонентам окружающей среды лежит в основе аллергических заболеваний, а к резус-антигену (возникает при резус-несовместимости беременности) - в основе гемолитической болезни новорожденных. Феномен иммунологической памяти используется в практике вакцинации людей.
Иммунологическая толерантность
Иммунологическая толерантность - явление, противоположное иммунному ответу и иммунологической памяти, проявляющееся в том, что на введение антигена вместо выработки иммунитета в организме развивается ареактивность, инертность, отсутствие ответа на антиген.
Иммунный ответ против собственных тканей организма в нормальных условиях не развивается, т.е. иммунная система толерантна к подавляющему большинству Аг тканей организма (аутоантигены). Искусственная толерантность к чужеродным Аг может быть вызвана иммунизацией по определенной схеме (например, толерантность «низкой дозы» - дробное введение Аг в возрастающих количествах или толерантность «высокой дозы» - однократное введение Аг в высокой дозе).
ВИДЫ ИММУНИТЕТА
Многообразие систем защиты организма позволяют человеку оставаться невосприимчивым к действию инфекционных агентов.
Видовой иммунитет (врожденный) - генетически закрепленная невосприимчивость присущая каждому виду. Например, человек никогда не заболевает чумой крупного рогатого скота. Крысы резистентны к дифтерийному токсину.
Приобретенный иммунитет формируется в течение жизни индивидуума и не передается по наследству; может быть естественным и искусственным, активным и пассивным.
Естественно приобретенный иммунитет (активный) развивается после перенесенного инфекционного заболевания, протекавшего в клинически выраженной форме, либо после скрытых контактов с микробными Аг (так называемая бытовая иммунизация). В зависимости от свойств возбудителя и состояния иммунной системы организма невосприимчивость может быть пожизненной (например, после кори), длительной (после брюшного тифа) или сравнительно кратковременной (после гриппа).
Инфекционный (нестерильный) иммунитет - особая форма приобретенной невосприимчивости; не является следствием перенесенной инфекции, обусловлен наличием инфекционного агента в организме. Невосприимчивость исчезает сразу после элиминации возбудителя из организма (например, туберкулез; вероятно, малярия).
Естественный пассивный иммунитетсвязан с переносом IgG от матери к плоду через плаценту (передача по вертикали) или с грудным молоком (SIgA) новорожденному. Это обеспечивает устойчивость новорожденного ко многим возбудителям в течение некоторого, обычно индивидуально варьирующего срока.

Рис. 3. Естественный пассивный иммунитет ребенка создается за счет материнских антител.
Искусственно приобретенный иммунитет. Состояние невосприимчивости развивается в результате вакцинации, серопрофилактики (введение сыворотки) и других манипуляций.
- Активно приобретенный иммунитет развивается после иммунизации ослабленными или убитыми микроорганизмами либо их антигенами. В обоих случаях организм активно участвует в создании невосприимчивости, отвечая развитием иммунного ответа и формированием пула клеток памяти.
- Пассивно приобретенный иммунитет достигается введением готовых АТ или, реже, сенсибилизированных лимфоцитов. В таких ситуациях иммунная система реагирует пассивно, не участвуя в своевременном развитии соответствующих иммунных реакций.
Иммунитет может формироваться против микроорганизмов, их токсинов, вирусов, антигенов опухолей. В этих случаях иммунитет называют антимикробным, антитоксическим, антивирусным, противоопухолевым соответственно. При трансплантации несовместимых тканей возникает трансплантационный иммунитет (реакция отторжения трансплантата).
Поступление в организм антигена через дыхательные пути, пищеварительный тракт и другие участки слизистых поверхностей и кожи нередко обуславливает развитие выраженной локальной иммунной реакции. В таких случаях речь идет о местном иммунитете.
Дата добавления: 2017-10-04; просмотров: 2149;
Поиск по сайту
Узнать еще
- Arthropoda. Клещи. Систематика. Морфология. Медицинское значение.
- Arthropoda. Паукообразные. Систематика. Географическое распространение. Морфология. Скорпионы. Пауки. Медицинское значение.
- Arthropoda..Систематика.Насекомые.Морфология.Классификация.Медицинское значение.
- Arthropoda.Систематика.Блохи.Виды блох.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское и эпидемиологическое значение.Меры борьбы.
- Arthropoda.Систематика.Мошки,мокрецы,слепни,оводы.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское значение,меры борьбы.
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- D) Система класифікації за підтримкою багато гілкового виконання програми.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
