Хемиосмотическая гипотеза Митчелла
Каким образом осуществляется сопряжение между окислением НАДН и фосфорилированием АДФ? Для осуществления окислительного фосфорилирования необходимо наличие целостной внутренней митохондриальной мембраны. Митохондрии содержатся в цитоплазме всех аэробных эукариотических клеток и располагаются вблизи от структур, нуждающихся в АТФ, который является продуктом их жизнедеятельности, или же поблизости от источников клеточного топлива, в котором они сами нуждаются. Митохондрия ограничена двумя отдельными мембранами. Внешняя мембрана гладкая, внутренняя имеет нерегулярно чередующиеся складки – кристы. Внутренняя часть митохондрий называется матриксом. Каждая из частей мембраны содержит специфические ферменты. Внешняя мембрана содержит ферменты метаболизма жирных кислот, фосфолипидов и липидов, ферменты, удлиняющие цепи жирных кислот. Приблизительно 20-25% общего белка внутренней мембраны составляют белки ферментов, участвующих в функционировании системы переноса электронов и окислительного фосфорилирования – это флавопротеиды, цитохромы и ферменты, ответственные за образование АТФ. Флавопротеиды и цитохромы располагаются на мембране в виде отдельных групп таким образом, что каждая из групп представляет собой самостоятельно функционирующую единицу, содержащую в определенных молярных соотношениях все необходимые ферменты. Такие группы называют дыхательными ансамблями. Дыхательные ансамбли распределены на плоскости мембран. Компоненты дыхательной цепи непосредственно связаны с фактором фосфорилирования F1 (фактор сопряжения). Ферменты цикла Кребса содержатся в матриксе.
Окислительное фосфорилирование является процессом, при котором выделение энергии при окислении субстрата сопряжено с синтезом АТФ. Для объяснения сопряжения окисления с фосфорилированием Митчеллом была выдвинута гипотеза. Он предположил, что сопряжение переноса электронов и синтеза АТФ обеспечивается протонным градиентом. Согласно этой модели, перенос электронов по дыхательной цепи приводит к выбросу протонов из матрикса на цитоплазматическую сторону внутренней митохондриальной мембраны, где возрастает концентрация протонов Н+. Возникает электрохимический градиент протонов в мембране. Этот градиент состоит из двух составных частей – разницы в концентрации водородных ионов (рН) и разницы в электрических потенциалах. Энергия этого градиента является движущей силой процесса синтеза АТФ, в ходе которого происходит обратное перемещение протонов по направлению градиента.
При переносе водорода на кислород каждая пара электронов, акцептируемых от субстрата, трижды пересекает мембрану туда и обратно и каждый раз выводит 2 протона. Первый переход осуществляется через восстановленный флавинмононуклеотид –QH2.
Общее число протонов, переносимых при восстановлении КоQ, равно 4. Каждая молекула QH2 отдает электрон цитохрому с1. Далее электрон по компонентам дыхательной цепи – цитохромам с и а передается терминальному цитохрому – а3, который окисляется молекулой кислорода. При этом каждый атом кислорода принимает 2 электрона и присоединяет 2 протона, образуя молекулу воды. Таким образом, каждая пара электронов, переносимая от НАДН к кислороду, приводит к перемещению шести протонов от внутренней к внешней поверхности мембраны. Этот процесс завершается на стадии АТФазы (комплекс F1–Fo), где каждые два перенесенных протона осуществляют синтез одной молекулы АТФ (рис.15).
Есть несколько гипотез о механизме синтеза АТФ комплексом (F1–Fo). Согласно Митчеллу, фосфатная группа связывается ферментом, активный центр которого находится в F1 части комплекса вблизи конца канала, по которому переносится протон. 2 протона переносятся по этому каналу под действием градиента рН и мембранного потенциала. Эти протоны атакуют один из кислородов фосфата, который отщепляется в виде молекулы воды. При этом фосфатная группа превращается в реакционноспособную частицу, способную реагировать с АДФ с образованием АТФ.
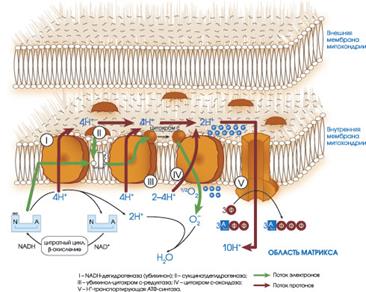
Рис.15. Дыхательная цепь митохондрий
Другая гипотеза заключается в том, что протоны вызывают изменения конформации белка вблизи активного центра, приводящие к синтезу АТФ. В активном центре такого фермента происходит самопроизвольное соединение АДФ и Фн. Образующаяся молекула АТФ остается связанной с ферментом и для ее отщепления необходимо затратить энергию. Энергия может быть получена в результате присоединения протона к ферменту с изменением его конформации.
Дата добавления: 2017-04-05; просмотров: 2438;
Поиск по сайту
Узнать еще
- Волновая природа материи. Гипотеза Луи-де-Бройля.
- Вопрос 1. Гипотеза де Бройля и ее экспериментальное подтверждение.
- ВТОРАЯ ГИПОТЕЗА ПРОЧНОСТИ - НАИБОЛЬШИХ ЛИНЕЙНЫХ ДЕФОРМАЦИЙ
- Гипотеза А.Н.Леонтьева
- Гипотеза Ампера. Молекулярные токи. Намагниченность. Диамагнетики и парамагнетики. Напряженность магнитного поля.
- Гипотеза глобального изменения климата Земли
- Гипотеза де Бройля. Волновые свойства частиц
- Гипотеза де Бройля. Корпускулярно волновой дуализм микрочасктиц.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
