Тема 14. ФИЗИОЛОГИЯ ЗРИТЕЛЬНОЙ СИСТЕМЫ.
Зрение в жизни человека имеет огромное значение. Это основной сенсорный канал, который связывает его с внешним миром. (Более 70% информации из окружающего мира человек получает через глаза.) Зрительная система человека устроена очень сложно.
Зрительная система (зрительный анализатор) включает следующие анатомические образования:
· периферический парный орган зрения — глаз;
· черепно-мозговые нервы: зрительный нерв — II-я пара, глазодвигательный нерв — III-я пара, блоковый нерв — IV-я пара и отводящий нерв — VI-я пара;
· латеральное коленчатое тело промежуточного мозга (с подкорковыми зрительными центрами);
· передние бугры четверохолмия среднего мозга (первичные зрительные центры);
· подушка таламуса,
· зрительная кора.
Схема зрительной системы приведена на рис.

Строение глаза. Глаз представляет собой сферический орган, покрытый плотной фиброзной оболочкой, склерой. Склера спереди переходит в прозрачную роговицу. Внутренняя поверхность склеры выстлана двумя тонкими оболочками - сосудистой и сетчаткой. Сосудистая оболочка, содержащая многочисленные сосуды, питающие глаз, расположена между склерой и сетчаткой. Впереди сосудистая оболочка переходит в ресничное тело (поясок) и далее в радужку.В радужке находятся гладкие мышечные волокна, степень напряжения которых определяет диаметр зрачка. Дно глазного бокала выстлано сетчаткой. Сетчатка - это слой, образованный нервными элементами: здесь расположены фоторецепторы и вставочные нейроны. Аксоны ганглиозных клеток сетчатки образуют зрительный нерв. Зрительная ось пересекает сетчатку в центральной ямке, небольшом углублении в сетчатке. Центральная ямка составляет область наибольшей остроты зрения.
Хрусталик делит глаз на два отсека с жидким содержимым: передняя камера заполнена водянистой влагой; позади хрусталика находится сосудистая масса - стекловидное тело.

Вращение глазного яблока в глазнице осуществляется тремя парами мышц, которые иннервируются глазодвигательными черепными нервами.
Нормальным раздражителем органа зрения является свет. Перед тем как попасть на сетчатку, свет должен пройти через прозрачную роговицу, водянистую влагу, зрачок, хрусталик и стекловидное тело. Оптика глаза создает на сетчатке уменьшенное, перевернутое изображение, а при помощи аккомодации это изображение становится резким.
Зрачковый рефлекс.Зрачком называют отверстие в центре радужной оболочки, через которое лучи света проходят внутрь глаза. Зрачок повышает четкость изображения на сетчатке, увеличивая глубину резкости глаза. Пропуская только центральные лучи, он улучшает изображение на сетчатке также за счет устранения сферической аберрации. Если прикрыть глаз от света, а затем открыть его, то расширившийся при затемнении зрачок быстро сужается («зрачковый рефлекс»). Мышцы радужной оболочки изменяют величину зрачка, регулируя поток света, попадающий в глаз. Так, на очень ярком свету зрачок имеет минимальный диаметр (1,8 мм), при средней дневной освещенности он расширяется (2,4 мм), а в темноте расширение максимально (7,5 мм). Это приводит к ухудшению качества изображения на сетчатке, но увеличивает чувствительность зрения. Предельное изменение диаметра зрачка изменяет его площадь примерно в 17 раз. Во столько же раз меняется при этом световой поток. Между интенсивностью освещения и диаметром зрачка имеется логарифмическая зависимость. Реакция зрачка на изменение освещенности имеет адаптивный характер, так как стабилизирует освещенность сетчатки.
В радужной оболочке имеется два вида мышечных волокон, окружающих зрачок: кольцевые, иннервируемые парасимпатическими волокнами глазодвигательного нерва, а также радиальные, иннервируемые симпатическими нервами. Первая состоит из окружающих отверстие циркулярных волокон, тогда как вторая образована радиальными волокнами, отходящими от зрачка подобно спицам колеса. Сокращение первых вызывает сужение, сокращение вторых — расширение зрачка. Зрачки расширяются во время боли, при гипоксии, а также при эмоциях, усиливающих возбуждение симпатической системы (страх, ярость). Расширение зрачков — важный симптом ряда патологических состояний, например болевого шока, гипоксии.
У здоровых людей размеры зрачков обоих глаз одинаковые. При освещении одного глаза зрачок другого тоже суживается; такая реакция называется содружественной. В некоторых патологических случаях размеры зрачков обоих глаз различны (анизокория).
Формирование изображения. На пути к светочувствительной оболочке глаза (сетчатке) лучи света проходят через несколько прозрачных сред — роговицу, водянистую влагу, хрусталик и стекловидное тело. Прохождение световых лучей через искривленную поверхность, разграничивающую две среды с различной оптической плотностью, сопровождается преломлением лучей, или рефракцией. При прохождении лучей через глаз они преломляются на четырех поверхностях раздела:
1) между воздухом и роговицей;
2) между роговицей и водянистой влагой;
3) между водянистой влагой и хрусталиком;
4) между хрусталиком и стекловидным телом.
Преломление зависит от угла падения световых лучей на поверхность линзы; чем больше угол падения, тем сильнее преломление луча. Лучи, проходящие через центр линзы перпендикулярно к ней, не преломляются вовсе.

В результате преломления на сетчатке получается изображение, резко уменьшенное и перевернутое вверх ногами и справа налево.
Аккомодация. Аккомодацией называют приспособление глаза к ясному видению объектов, удаленных на разное расстояние. Для ясного видения объекта необходимо, чтобы он был сфокусирован на сетчатке, т. е. чтобы лучи от всех точек его поверхности проецировались на поверхность сетчатки. Когда мы смотрим на далекие предметы, их изображение сфокусировано на сетчатке и они видны ясно. Зато изображение близких предметов при этом расплывчато, так как лучи от них собираются за сетчаткой. Главную роль в аккомодации играет хрусталик, изменяющий свою кривизну и, следовательно, преломляющую способность. При рассматривании близких предметов хрусталик делается более выпуклым, благодаря чему лучи, расходящиеся от какой-либо точки объекта, сходятся на сетчатке. Механизмом аккомодации является сокращение ресничных мышц, которые изменяют выпуклость хрусталика. Хрусталик заключен в тонкую прозрачную капсулу, которую всегда растягивают, т. е. уплощают, волокна ресничного пояска (циннова связка). Сокращение гладких мышечных клеток ресничного тела уменьшает тягу цинновых связок, что увеличивает выпуклость хрусталика в силу его эластичности. Ресничные мышцы иннервируются парасимпатическими волокнами глазодвигательного нерва. Введение в глаз атропина вызывает нарушение передачи возбуждения к этой мышце, ограничивает аккомодацию глаза при рассматривании близких предметов. Наоборот, парасимпатомиметические вещества — вызывают сокращение этой мышцы.
Для нормального глаза молодого человека дальняя точка ясного видения лежит в бесконечности. Далекие предметы он рассматривает без всякого напряжения аккомодации, т. е. без сокращения ресничной мышцы. Ближайшая точка ясного видения находится на расстоянии 10 см от глаза.
Острота зрения отражает способность оптической системы глаза строить четкое изображение на сетчатке. Она измеряется путем определения наименьшего расстояния между двумя точками, при котором их изображения не сливаются. Это расстояние должно быть достаточным для того, чтобы лучи от обеих точек попадали на разные рецепторы сетчатки. Остроту зрения можно проверить с помощью таблиц Снеллена. У многих людей изображение на сетчатке всегда получается нечетким. Это бывает связано либо с необычной формой глазного яблока, либо с неправильной кривизной роговицы или хрусталика.
Старческая дальнозоркость. Хрусталик с возрастом теряет эластичность, и при изменении натяжения цинновых связок его кривизна меняется мало. Поэтому ближайшая точка ясного видения находится теперь не на расстоянии 10 см от глаза, а отодвигается от него. Близкие предметы при этом видны плохо. Это состояние называется старческой дальнозоркостью, или пресбиопией. Пожилые люди вынуждены пользоваться очками с двояковыпуклыми линзами.
Аномалии рефракции глаза. Две главные аномалии рефракции глаза — близорукость, или миопия, и дальнозоркость, или гиперметропия, — обусловлены не недостаточностью преломляющих сред глаза, а изменением длины глазного яблока.
Близорукость. Если продольная ось глаза слишком длинная, то лучи от далекого объекта сфокусируются не на сетчатке, а перед ней, в стекловидном теле. Такой глаз называется близоруким, или миопическим. Чтобы ясно видеть вдаль, необходимо перед близорукими глазами поместить вогнутые стекла, которые отодвинут сфокусированное изображение на сетчатку.

Рис. Близорукость(I, II) и дальнозоркость(III,IV)
Дальнозоркость. Противоположна близорукости дальнозоркость, или гиперметропия. В дальнозорком глазу продольная ось глаза укорочена, и поэтому лучи от далекого объекта фокусируются не на сетчатке, а за ней. Этот недостаток рефракции может быть компенсирован аккомодационным усилием, т. е. увеличением выпуклости хрусталика. Поэтому дальнозоркий человек напрягает аккомодационную мышцу, рассматривая не только близкие, но и далекие объекты. При рассматривании близких объектов аккомодационные усилия дальнозорких людей недостаточны.
Поэтому для чтения дальнозоркие люди должны надевать очки с двояковыпуклыми линзами, усиливающими преломление света.
Гиперметропию не следует путать со старческой дальнозоркостью. Общее у них лишь то, что необходимо пользоваться очками с двояковыпуклыми линзами.
Астигматизм. К аномалиям рефракции относится также астигматизм, т. е. неодинаковое преломление лучей в разных направлениях (например, по горизонтальному и вертикальному меридиану). Астигматизм обусловлен не строго сферической поверхностью роговой оболочки. При астигматизме сильных степеней эта поверхность может приближаться к цилиндрической, что исправляется цилиндрическими очковыми стеклами, компенсирующими недостатки роговицы.
Структура и функции сетчатки. Сетчатка представляет собой внутреннюю светочувствительную оболочку глаза. По своему строению и происхождению представляет собой нервный центр, в котором происходят первичная обработка зрительных сигналов, преобразование их в нервные импульсы, передающиеся в головной мозг.
Сетчатка состоит из четырех слоев клеток: пигментного слоя, слоя фоторецепторов и двух слоев нейронов сетчатки. Наружный (ближайший к склере) слой образован пигментными клетками. Слой фоторецепторов расположен между пигментными и нервными клетками. Во внутреннем (прилегающем к стекловидному телу) слое находятся ганглиозные нервные клетки, аксоны которых образуют зрительный нерв.

Рис. Строение сетчатки глаза
Вследствие такого «обратного» расположения слоев сетчатки позвоночных животных свет у них, прежде чем попасть на фоторецепторы, должен пройти оба слоя нервных клеток; по мере прохождения через эти слои многие лучи рассеиваются на нейронах. Из-за этого рассеивания качество изображения на сетчатке страдает.
Лишь в небольшом участке сетчатки - центральной ямке — формируется четкое изображение. Здесь слои нейронов смещены к периферии и фоторецепторы открыты для восприятия световых лучей. Свет непосредственно падает на рецепторы центральной ямки, не рассеиваясь на нейронах. В этом участие содержится большое количество маленьких фоторецепторов в виде колбочек, что повышает остроту зрения.
Рассмотрим структуру и функции слоев сетчатки, следуя от наружного (заднего, наиболее удаленного от зрачка) слоя сетчатки к внутреннему (расположенному ближе к зрачку) ее слою.
Пигментный слой.Этот слой образован одним рядом эпителиальных клеток, содержащих большое количество различных внутриклеточных органелл, включая меланосомы, придающие этому слою черный цвет. Пигмент - фуксин, называемый также экранирующим пигментом, поглощает доходящий до него свет, препятствуя тем самым его отражению и рассеиванию, что способствует четкости зрительного восприятия. Клетки пигментного эпителия имеют многочисленные отростки, которые плотно окружают светочувствительные наружные сегменты палочек и колбочек, Пигментный эпителий играет решающую роль в целом ряде функций, в том числе в ресинтезе (регенерации) зрительного пигмента после его обесцвечивания, в фагоцитозе и переваривании обломков наружных сегментов палочек и колбочек, иными словами, в механизме постоянного обновления наружных сегментов зрительных клеток, в защите зрительных клеток от опасности светового повреждения, а также в переносе к фоторецепторам кислорода и других необходимых им веществ. Следует отметить, что контакт между клетками пигментного эпителия и фоторецепторами достаточно слабый. Именно в этом месте происходит отслойка сетчатки — опасное заболевание глаз. Отслойка сетчатки приводит к нарушению зрения не только вследствие ее смещения с места оптического фокусирования изображения, но и вследствие дегенерации рецепторов из-за нарушения контакта с пигментным эпителием, что приводит к серьезнейшему нарушению метаболизма самих рецепторов. Метаболические нарушения усугубляются тем, что нарушается доставка питательных веществ из капилляров сосудистой оболочки глаза, а сам слой фоторецепторов капилляров не содержит (аваскуляризован).
Фоторецепторы. К пигментному слою изнутри примыкает слой фоторецепторов. В сетчатке имеется два вида фоторецепторов - палочки и колбочки. Палочки, чувствительность которых выше при слабом освещении, ответственны за «сумеречное зрение»; колбочки - воспринимают различные цвета и отвечают за «дневное зрение». В сетчатке каждого глаза человека находится 6—7 млн колбочек и 110—123 млн палочек. Они распределены в сетчатке неравномерно. Колбочки сосредоточены преимущественно в центре сетчатки, в частности в центральной ямке (до 140 тыс. на 1 мм2). По направлению к периферии сетчатки их число уменьшается, а число палочек возрастает, так что на дальней периферии имеются только палочки.
Цвет воспринимается лучше всего при действии света на центральную ямку сетчатки, где расположены почти исключительно колбочки. Здесь же и наибольшая острота зрения. По мере удаления от центра сетчатки восприятие цвета и пространственное разрешение становятся все хуже. Периферия сетчатки, где находятся исключительно палочки, не воспринимает цвета. Зато световая чувствительность колбочкового аппарата сетчатки во много раз меньше, чем палочкового, поэтому в сумерках из-за резкого понижения «колбочкового» зрения и преобладания «периферического» зрения мы не различаем цвет.
Нарушение функции палочек, возникающее при недостатке в пище витамина А, вызывает расстройство сумеречного зрения — так называемую куриную слепоту: человек совершенно слепнет в сумерках, но днем зрение остается нормальным. Наоборот, при поражении колбочек возникает "светобоязнь": человек видит при слабом свете, но слепнет при ярком освещении. В этом случае может развиться и полная цветовая слепота — ахромазия.
Строение фоторецепторной клетки. Фоторецепторная клетка — палочка или колбочка — состоит из чувствительного к действию света наружного сегмента, содержащего зрительный пигмент, внутреннего сегмента и соединительной ножки. Палочка и колбочка сетчатки обращены своими светочувствительными наружными сегментами к пигментному эпителию, т. е. в сторону, противоположную свету. У человека наружный сегмент фоторецептора (палочка или колбочка) содержит около тысячи фоторецепторных дисков. Наружный сегмент палочки намного длиннее, чем колбочки, и содержит больше зрительного пигмента. Это частично объясняет более высокую чувствительность палочки к свету: палочку может возбудить всего один квант света, а для активации колбочки требуется больше сотни квантов. Фоторецепторный диск образован двумя мембранами, соединенными по краям.
Внутренний сегмент фоторецептора соединен с наружным сегментом модифицированной ресничкой, которая содержит девять пар микротрубочек. Внутренний сегмент содержит крупное ядро и весь метаболический аппарат клетки, в том числе митохондрии, обеспечивающие энергетические потребности фоторецептора, и систему белкового синтеза, обеспечивающую обновление мембран наружного сегмента. Здесь происходят синтез и включение молекул зрительного пигмента в фоторецепторную мембрану диска. За час на границе внутреннего и наружного сегмента в среднем заново образуется три новых диска. Затем они медленно (у человека примерно в течение 2—3 нед) перемещаются от основания наружного сегмента палочки к его верхушке, В конце концов верхушка наружного сегмента, содержащая до сотни теперь уже старых дисков, обламывается и фагоцитируется клетками пигментного слоя. Это один из важнейших механизмов защиты фоторецепторных клеток от накапливающихся в течение их световой жизни молекулярных дефектов.
Наружные сегменты колбочек также постоянно обновляются, но с меньшей скоростью. Интересно, что существует суточный ритм обновления: верхушки наружных сегментов палочек в основном обламываются и фагоцитируются в утреннее и дневное время, а колбочек — в вечернее и ночное.
Зрительные пигменты. В палочках сетчатки человека содержится пигмент родопсин, или зрительный пурпур, максимум спектра поглощения которого находится в области 500 нанометров (нм). В наружных сегментах трех типов колбочек (сине-, зелено-и красно-чувствительных) содержится три типа зрительных пигментов, максимумы спектров поглощения которых находятся в синей (420 нм), зеленой (531 нм) и красной (558 нм) частях спектра. Красный колбочковый пигмент получил название «йодопсин». Молекула зрительного пигмента сравнительно небольшая (с молекулярной массой около 40 килодальтон), состоит из большей белковой части (опсина) и меньшей хромофорной (ретиналь, или альдегид витамина А).
Ретиналь может находиться в различных пространственных конфигурациях, т. е. изомерных формах, но только одна из них — 11-цис-изомер ретиналя выступает в качестве хромофорной группы всех известных зрительных пигментов. Источником ретиналя в организме служат каротиноиды, поэтому недостаток их приводит к дефициту витамина А и, как следствие, к недостаточному ресинтезу родопсина, что в свою очередь является причиной нарушения сумеречного зрения, или «куриной слепоты».
Молекулярная физиология фоторецепции. На первом этапе зрительного восприятия световая энергия трансформируется в нервное возбуждение. Этот процесс происходит в наружных члениках фоторецепторов, содержащих фотопигмент: в колбочках – йодопсин, в палочках – родопсин. Молекула фотопигмента состоит из белковой части – опсина и части, поглощающей свет – ретиналя. Колбочки и палочки отличаются друг от друга по структуре и зрительным пигментам, а также по некоторым особенностям функционирования. Однако основные принципы возбуждения у них одинаковы. Внутриклеточные регистрации от колбочек и палочек животных показали, что в темноте вдоль фоторецептора течет темновой ток, выходящий из внутреннего сегмента и входящий в наружный сегмент.
В темноте фоторецептор непрерывно выделяет медиатор, который действует деполяризующим образом на мембраны постсинаптических отростков горизонтальных и биполярных клеток. Освещение приводит к блокаде тока.
При поглощении кванта света молекулой зрительного пигмента (родопсина) в ней происходит мгновенная изомеризация ее хромофорной группы: 11-цис-ретиналь выпрямляется и превращается в транс-ретиналь. Свет выполняет роль спускового, или триггерного, фактора, запускающего механизм фоторецепции. Вслед за фотоизомеризацией ретиналя происходят пространственные изменения в белковой части молекулы. Результатом всех событий, вызванных поглощением кванта света, становится закрытие ионных каналов в плазматической мембране наружного сегмента, которые были открыты в темноте и через которые внутрь клетки входили Na+ и Са2+.
Уменьшение или прекращение входа внутрь наружного сегмента Na+ приводит к гиперполяризации клеточной мембраны, т. е. возникновению на ней рецепторного потенциала.
Гиперполяризационный рецепторный потенциал, возникший на мембране наружного сегмента, распространяется затем вдоль клетки до ее пресинаптического окончания и приводит к уменьшению скорости выделения медиатора (глутамата). Таким образом, фоторецепторный процесс завершается уменьшением скорости выделения нейромедиатора из пресинаптического окончания фоторецептора.
Не менее сложен и совершенен механизм восстановления исходного темнового состояния фоторецептора, т. е. его способности ответить на следующий световой стимул. Для этого необходимо вновь открыть ионные каналы в плазматической мембране. Через открытый канал внутрь клетки вновь начинают входить Na+ и Са2+, деполяризуя мембрану рецептора и переводя его в «темновое» состояние. Из пресинаптического окончания деполяризованного рецептора вновь ускоряется выход медиатора.
Нейроны сетчатки.
Фоторецепторы сетчатки синаптически связаны с биполярными нейронами. При действии света уменьшается выделение медиатора (глутамата) из фоторецептора, что приводит к гиперполяризации мембраны биполярного нейрона. От него нервный сигнал передается на ганглиозные клетки, аксоны которых являются волокнами зрительного нерва. Передача сигнала как с фоторецептора на биполярный нейрон, так и от него на ганглиозную клетку происходит безымпульсным путем. Биполярный нейрон не генерирует импульсов ввиду предельно малого расстояния, на которое он передает сигнал.
На 130 млн фоторецепторных клеток приходится только 1 млн 250 тыс. ганглиозных клеток, аксоны которых образуют зрительный нерв. Это значит, что импульсы от многих фоторецепторов сходятся (конвергируют) через биполярные нейроны к одной ганглиозной клетке. Фоторецепторы, соединенные с одной ганглиозной клеткой, образуют рецептивное поле ганглиозной клетки. Рецептивные поля различных ганглиозных клеток частично перекрывают друг друга. Таким образом, каждая ганглиозная клетка суммирует возбуждение, возникающее в большом числе фоторецепторов. Это повышает световую чувствительность, но ухудшает пространственное разрешение. Лишь в центре сетчатки, в районе центральной ямки, каждая колбочка соединена с одной так называемой карликовой биполярной клеткой, с которой соединена также всего одна ганглиозная клетка. Это обеспечивает здесь высокое пространственное разрешение, но резко уменьшает световую чувствительность.
Таким образом, сигналы передаются через цепочку из трех основных типов клеток:
1) фоторецепторы (палочки и колбочки);
2) биполярные клетки;
3) ганглиозные клетки.
Взаимодействие соседних нейронов сетчатки обеспечивается горизонтальными и амакриновыми клетками. Горизонтальные клетки меняют синаптическую передачу на уровне переключения от фоторецепторов к биполярам, амакриновые клетки - на уровне переключения от биполяров к ганглиозным клеткам. Амакриновые клетки осуществляют боковое торможение между соседними ганглиозными клетками. На амакриновых клетках оканчиваются центробежные волокна, идущие из головного мозга. Амакриновые клетки управляются от биполяров через деполяризующие синапсы: увеличение деполяризующего синаптического тока вызывает деполяризацию амакриновой клетки, а уменьшение – гиперполяризацию. Горизонтально ветвящиеся отростки амакриновых клеток могут интегрировать влияния от большого числа биполярных клеток. Выходной сигнал, возникающий от такого интегрирования, влияет на активность ганглиозных клеток сетчатки.
Место выхода зрительного нерва из глазного яблока — диск зрительного нерва, называют слепым пятном. Оно не содержит фоторецепторов и поэтому нечувствительно к свету. Мы не ощущаем наличия «дыры» в сетчатке.
Выходными элементами сетчатки являются ганглиозные клетки. Большинство ганглиозных клеток сетчатки имеют концентрические рецептивные поля: при освещении одной из зон поля ганглиозная клетка возбуждается (on‑эффект), а при ее затемнении тормозится. Взаимоотношения могут быть обратными. В соответствии с этим различают рецептивные поля с on‑центром (возбуждаются при освещении центра) и с off‑центром (возбуждаются при его затемнении).
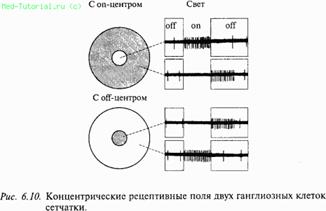
Центральную зону рецептивного поля окружает концентрическая тормозная зона. Нейроны с наименьшим размером центральной зоны локализуются в центральной части сетчатки. Тормозная кайма вокруг центральной зоны рецептивного поля не обнаруживается при низком уровне освещенности. В настоящее время предполагают, что тормозная кайма образуется горизонтальными клетками сетчатки по механизму латерального торможения, т.е. чем сильнее возбужден центр рецептивного поля, тем большее тормозное влияние он оказывает на периферию. Благодаря двум типам ганглиозных клеток с on‑ и off‑центрами обнаружение как светлых, так и темных объектов в поле зрения обеспечивается уже на уровне сетчатки.
Как показали исследования на животных с цветовым зрением, у них существует цветооппонентная организация рецептивных полей ганглиозных клеток сетчатки, т.е. данная ганглиозная клетка получает возбуждающие и тормозные входы от колбочек, имеющих разную спектральную чувствительность. Например, если «красные» колбочки оказывают возбуждающее действие на данную ганглиозную клетку, то «синие» колбочки ее затормаживают. Обнаружены различные комбинации возбуждающих и тормозящих входов от разных классов колбочек (разных цветоприемников). Благодаря такой организации рецептивные поля отдельных ганглиозных клеток становятся избирательными к освещению определенного спектрального состава. Например, у человека, имеющего трихроматическое зрение, значительная часть цветооппонентных ганглиозных клеток связана в разных вариантах со всеми тремя цветоприемниками: возбуждение от красночувствительного приемника и торможение от сине‑ и зеленочувствительных приемников; возбуждение от синечувствительных и торможение от зелено‑ и красночувствительных и т.д.
Световая и темновая адаптация сетчатки является многокомпонентным процессом и складывается из целого ряда последовательных реакций. Самые периферические процессы – изменение диаметра зрачка и ретиномоторный эффект. Ретиномоторный эффект у позвоночных состоит в том, что от яркого света экранируются только палочки за счет движения меланина по отросткам пигментного эпителия между наружными сегментами фоторецепторов. Значительно меньше исследованы механизмы адаптации на уровне самих рецепторов и соответствующих синапсов. В них, по‑видимому, принимают участие ионы кальция, а также имеет место изменение переходных процессов в горизонтальных клетках.
Анализ зрительных сигналов нейронами наружного коленчатого тела. Следующий уровень обработки зрительной информации находится в таламусе – это наружное коленчатое тело.
В наружное коленчатое тело каждой стороны мозга поступают волокна от сетчаток обоих глаз. Поскольку зрительные волокна по пути к наружному коленчатому телу перекрещиваются (зрительный перекрест), к наружному коленчатому телу приходят волокна только от половины сетчатки каждого глаза: от темпоральной половины ипсилатеральной сетчатки и от назальной половины контралатеральной сетчатки.
В наружном коленчатом теле человека морфологически определено шесть слоев. В различные слои наружного коленчатого тела приходят разные волокна сетчатки. Топическая проекция поля зрения имеется в каждом из слоев коленчатого тела. Хотя в разные слои наружного коленчатого тела приходят ретинальные волокна от разных сетчаток (контра‑ и ипсилатеральной), эти проекции расположены одна под другой, так что можно выделить колонкообразный участок, пересекающий все слои коленчатого тела, который соответствует проекции одной точки поля зрения. При этом наиболее подробно в коленчатом теле представлена проекция центральной части поля зрения (на сетчатке соответствует центральной ямке).
Рецептивные поля этих нейронов также круглые, но меньшего размера, чем в сетчатке. Так же как и рецептивные поля ганглиозных клеток, все нейроны наружного коленчатого тела можно разделить на два класса: с on‑центром (освещение центра рецептивного поля активирует нейрон) и с off‑центром (нейрон активируется затемнением центра). В наружном коленчатом теле имеется примерно равное количество нейронов с on‑ и off‑центрами. Ответы нейронов, генерируемые в ответ на вспышку света, здесь короче, чем в сетчатке. Сходное строение рецептивных полей этих двух уровней зрительного анализатора (сетчатки и коленчатого тела) дает основание предполагать, что в структуре рецептивных полей нейронов наружного коленчатого тела отражаются свойства рецептивных полей ганглиозных клеток сетчатки. Подобно нейронам сетчатки, нейроны наружного коленчатого тела обладают цветовой чувствительностью.
Наружное коленчатое тело – это первый уровень, на котором вследствие неполного перекреста зрительных волокон происходит конвергенция от двух сетчаток. Конвергенция является необходимым условием стереоскопического восприятия трехмерного мира.
На уровне наружных коленчатых тел происходит взаимодействие афферентных сигналов, пришедших из сетчатки, с эфферентными сигналами из зрительной области коры, а также через ретикулярную формацию от слуховой и других сенсорных систем. Эти взаимодействия обеспечивают выделение наиболее существенных компонентов сенсорного сигнала и процессы избирательного зрительного внимания.
Анализ зрительных стимулов нейронами коры больших полушарий. Импульсные разряды нейронов наружного коленчатого тела по их аксонам поступают в затылочную часть полушарий большого мозга, где расположена первичная проекционная область зрительной зоны коры (поле 17).
В коре происходит значительно более специализированная и сложная, чем в сетчатке и в наружных коленчатых телах, переработка информации. В отличие от наружного коленчатого тела основная масса нейронов зрительных областей коры больших полушарий (поля 17, 18 и 19) специализирована на выделении ориентированных линий и контуров, которые составляют основные элементы зрительных стимулов. Указанная способность полностью обусловлена строением рецептивных полей корковых нейронов. В отличие от концентрических рецептивных полей предшествующих уровней зрительного анализатора (сетчатка и наружное коленчатое тело) рецептивные поля корковых нейронов имеют вытянутые антагонистические зоны, которые определенным образом ориентированы в поле зрения. Благодаря этому они способны выделять из цельного изображения отдельные фрагменты линий с той или иной ориентацией и расположением (детекторы ориентации) и избирательно на них реагировать.
Хьюбел и Визель подразделили рецептивные поля корковых нейронов зрительной системы на «простые», «сложные» и «сверхсложные». «Простые» рецептивные поля имеют удлинённую форму, к примеру, с центральной эллипсовидной зоной возбуждения и антагонистической зоной торможения по периферии эллипса. Либо они могут быть почти прямоугольными; при этом одна из длинных сторон прямоугольника является зоной возбуждения, а другая — антагонистической зоной торможения. Изображения, активирующие нейроны этих рецептивных полей, должны быть ориентированы определённым образом. Чтобы возбудить нейрон со «сложным» рецептивных полем, световому стимулу в виде полоски недостаточно быть правильно ориентированным — нужно ещё и двигаться, причём в строго определённом направлении. Для активации корковых нейронов со «сверхсложными» рецептивными полями зрительному стимулу в виде полоски необходимо обладать всеми вышеперечисленными свойствами, и к тому же длина этой полоски должна быть строго определённой.

В каждом небольшом участке зрительной зоны коры по ее глубине сконцентрированы нейроны с одинаковой ориентацией и локализацией рецептивных полей в поле зрения. Они образуют колонку нейронов, проходящую вертикально через все слои коры. Колонка — пример функционального объединения корковых нейронов, осуществляющих сходную функцию. Колонки объединяются в гиперколонки. Под гиперколонкой понимается участок коры, включающий весь набор ориентированных колонок (с ориентациями в пределах 180° и перекрывающиеся с ними две окулодоминантные колонки). Такая гиперколонка обрабатывает информацию от определенного участка сетчатки; информация от соседних участков обрабатывается такими же соседними гиперколонками. Однако из выше изложенного не следует, что нейроны, составляющие колонку, в функциональном отношении совершенно однотипны. Например, в зрительной коре кошки одна колонка содержит нейроны с простыми и сложными рецептивными полями. В этом случае общим признаком будет только ориентация рецептивных полей отдельных нейронов. В одной колонке могут находиться нейроны с одинаковой цветоизбирательностью и другими однотипными свойствами.
Как показывают результаты исследований последних лет, функциональное объединение отдаленных друг от друга нейронов зрительной зоны коры может происходить также за счет синхронности их разрядов. Многие нейроны зрительной зоны коры избирательно реагируют на определенные направления движения (дирекциональные детекторы) либо на какой-то цвет, а часть нейронов лучше всего отвечает на относительную удаленность объекта от глаз. Информация о разных признаках зрительных объектов (форма, цвет, движение) обрабатывается параллельно в разных частях зрительной зоны коры большого мозга.
Основой бинокулярного стереоскопического зрения является диспаратность – оценка различия проекций изображений на сетчатках обоих глаз. Известно, что 84% нейронов зрительной коры бинокулярны. Они реагируют при одновременной стимуляции двух сетчаток, при этом нейроны одной колонки имеют близкие значения диспаратности. Благодаря тому, что правый и левый глаза рассматривают один и тот же объект с разных точек, изображения объекта на правой и левой сетчатке сдвинуты относительно друг друга. Это различие изображений зависит от того, насколько приближен или отдален объект относительно точки пересечения оптических осей двух глаз (точка фиксации). Нейрофизиологической основой бинокулярного зрения является взаимодействие двух рецептивных полей, которые имеются у каждого бинокулярного нейрона зрительной коры (на контра‑ и ипсилатеральной сетчатке). Каждый бинокулярный нейрон избирателен к какой‑то одной определенной диспаратности (это определяется диспаратностью его рецептивных полей), а поскольку диспаратность зависит от удаленности объекта, реакции бинокулярных нейронов оказываются избирательными к определенной удаленности. В коре имеется целый набор нейронов с разной диспаратностью. Эта совокупность нейронов составляет механизм, измеряющий удаленность объекта. Описанный выше механизм диспаратности лежит в основе стереоскопического зрения.
Цветооппонентные нейроны в зрительной коре обезьян. Значительная часть цветооппонентных нейронов зрительной коры приматов имеет простые или концентрические рецептивные поля. Большинство нейронов с цветооппонентными рецептивными полями встречаются
Дата добавления: 2017-01-16; просмотров: 9313;
Поиск по сайту
Узнать еще
- Arthropoda. Клещи. Систематика. Морфология. Медицинское значение.
- Arthropoda. Паукообразные. Систематика. Географическое распространение. Морфология. Скорпионы. Пауки. Медицинское значение.
- Arthropoda..Систематика.Насекомые.Морфология.Классификация.Медицинское значение.
- Arthropoda.Систематика.Блохи.Виды блох.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское и эпидемиологическое значение.Меры борьбы.
- Arthropoda.Систематика.Мошки,мокрецы,слепни,оводы.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское значение,меры борьбы.
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- D) Система класифікації за підтримкою багато гілкового виконання програми.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
