Изучение структурных компонентов клеток и органов иммунной системы
ОБЩАЯ ХАРАКТЕРИСТИКА Т- И В-ЛИМФОЦИТОВ.
В организме человека содержится 75% Т-лимфоцитов, 15% В-лимфоцитов и 10% лимфоцитов, не несущих маркеров ни Т-, ни В-лимфоцитов. Маркерами, позволяющими дифференцировать Т- и В-лимфоциты, являются иммуноглобулиновые рецепторы, находящиеся на поверхности В-лимфоцитов, и рецепторы к бараньим эритроцитам, которыми снабжены Т-лимфоциты. Первые определяют с помощью иммунофлюоресцентного метода, вторые — методом Е-розеток. Более современный метод идентификации Т- и В-лимфоцитов основан на выявлении специфических поверхностных антигенов.
В-лимфоциты. Основной функцией В-лимфоцитов является синтез иммуно-глобулинов, который начинается после их созревания в плазматических клетках.
Связывание эпитопов антигена с гомологичными рецепторами на мембране В-лимфоцита является стимулом для его пролиферации, в результате которой образуется иммунологически однородный клон клеток, снабженных идентичными иммуноглобулиновыми рецепторами. Далее под влиянием клеточных медиаторов происходит дифференцировка клеток пролиферирующего клона с образованием клеток иммунологической памяти и клеток, продуцирующих антитела. Первые надолго сохраняются в организме, сохраняя память о данном антигене, т. е. способность быстро его распознать. Клетки иммунологической памяти обеспечивают более сильный и быстрый вторичный иммунный ответ при повторной встрече с тем же антигеном, например при повторной инфекции или вакцинации.
Клетки, продуцирующие антитела, увеличиваются в размерах, прекра-щают пролиферировать и начинают синтезировать антитела. Они получили название плазматических клеток, продолжительность жизни которых огра-ничивается несколькими днями. Однако за этот короткий срок плазмати-ческие клетки успевают синтезировать большое количество специфических к данному антигену антител, принадлежащих к разным классам иммуноглобулинов.
Т-лимфоциты в процессе дифференцировки и пролиферации образуют четыре субпопуляции, отличающиеся друг от друга по своим функциям. Две из них выполняют регуляторные функции. Это Т - хелперы, или помощники, и Т- супрессоры, или ингибиторы. Т-хелперы узнают детерминантные группы антигена на мембране макрофага и активируют при помощи медиаторов В-лимфоциты и эффекторные субпопуляции Т-лимфоцитов.
Т-супрессоры угнетают Т-хелперы, В-лимфоциты или плазматические клетки, что в конечном итоге задерживает синтез антител.
К субпопуляциям Т-лимфоцитов, выполняющим эффекторные функции, относятся цитотоксические клетки Т-киллеры — их основная функция уничтожать клетки-«мишени», несущие соответствующий чужеродный антиген, и Т-эффекторы, продуцирующие лимфокины. Т-эффекторы обеспечивают клеточный специфический иммунитет, непосредственно участвуют в формировании реакции гиперчувствительности замедленного типа (ГЗТ).
Рецепторы В- и Т-лимфоцитов наряду с антителами являются антигенрас-познающими молекулами иммунной системы. Они обладают способностью узнавать только одну определенную молекулярную структуру детерми-нантной группы антигена (эпитопа).
Рецепторы В-лимфоцитов представляют собой антигенсвязывающие участки молекул иммуноглобулинов, которые синтезируются данным В-лимфоцитом и частично остаются в составе его мембраны. Они фиксируются при помощи так называемого «якорного» сегмента иммуноглобулина (Fс-конец).
Рецепторы Т-лимфоцитов структурно похожи на иммуноглобулин. Это белки, состоящие из двух субъединиц — ά и β-расположенных на поверх-ности Т-лимфоцита. Каждая субъединица представляет собой полипеп-тидную цепь.
Антигенсвязывающий участок в молекуле рецептора Т-лимфоцита образован гипервариабельными участками полипептидных цепей, напоминающими аналогичные участки иммуноглобулинов, между которыми образуется полость.
Рис. 1. Возможные формы взаимодействия между Т- и В-клетками.
Установление того факта, что Т-клетки (Т) распознают главным образом несущуючасть, а В-клетки (В)— гаптенную часть антигена (АГ), позволило схематизировать возможные отношения между двумя типами клеток, возникающие в индукционный период развития гуморального иммунного ответа.
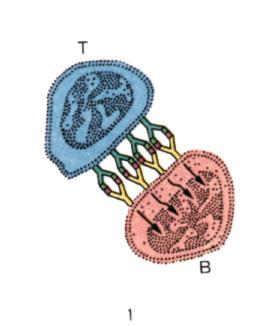
| 1. Концентрация АГ на поверхности Т-клеток включает в иммунный ответ В-клетки в результате множественного взаимодействия антигенраспоз-нающих рецепторов этих клеток с гаптенной частью АГ. |

| 2. В-клетки включаются в ответ после получения двойного сиг-нала: специфическо го — в рез-ультате взаимодействия анти-генраспознающих рецепторов В-клеток с гаптенной частью АГ, ассоциированного с Т-клеткой, и неспецифического, возникающего в результате вза-имодействия поверхностных структур Т- и В-клеток, не имеющих отношения к специи-фическому антигенному распознаванию. |

| 3. В-клетка получает двойной сигнал: первый в результате взаимодействия антигенраспоз-нающего рецептора В-клетки с гаптеном и второй — от комплекса АГ — антитело. |

| 4. В-клетка получает двойной сигнал: первый, специфический, который возникает, как и в пре-дыдущих случаях, при взаимо-действии В-клетки с гаптеном, и второй — неспецифический в виде продукции лимфокинов Т-клетками |
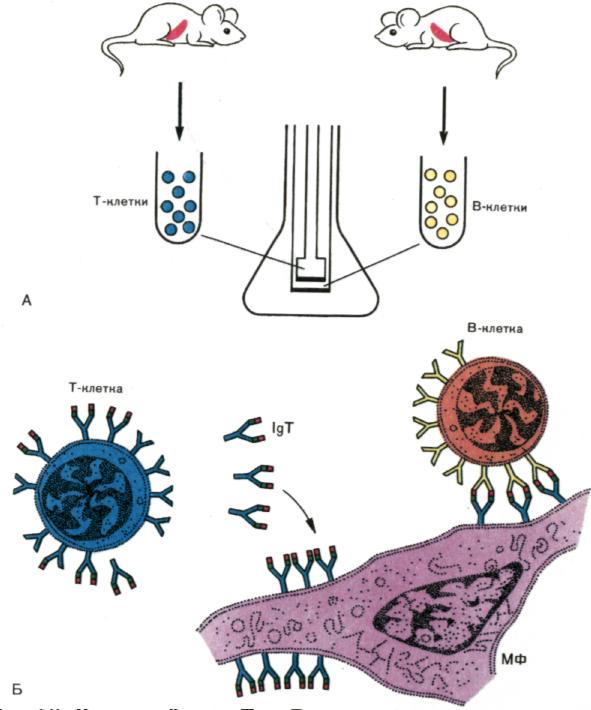
рис. 2. .Взаимодействие Т- и В-клеток без прямого контакта.
А. Т-клетки, примированные к конъюгату DNP-гемоцианин, помещали в верхний сосуд двухкамерной культуральной системы. В-клетки (клетки селезенки от мышей, примированных к DNP -флагеллину и обработанные анти-Тhу-1-сывороткой) вносили в нижний сосуд. Два сосуда разделены миллипоровой мембраной, которая препятствует клеточному обмену, но позволяет проникать макромолекулам.
Показано, что для развития иммунного ответа к DNP между Т- и В-клетками контакт не обязателен. В то же время требуется прямая физическая связь между макрофагом (МФ) и В-клеткой. Реализация иммунного ответа осуществляется гуморальным фактором Т-клеток, включающим иммуноглобулин, названный IgТ, и антиген.
Б. Предполагаемый механизм индукции В-клеток включает в качестве основного участника антигенспецифический фактор Т-клеток (IgТ, ассоциированный с антигеном). Фактор после выхода из Т-клетки сорбируется на МФ, создавая «батарею» антигенных детерминант, которые обеспечивают полноценное включение в ответ В-клеток.
Классификации. Ранее лимфоциты подразделяли на различные категории по их величине, значительно важнее их функциональная классификация.
(1). Величина клетки. Популяция лимфоцитов по этому признаку неоднородна, их размер в крови варьирует от 4,5 до 10 мкм: малые (4,5-6 мкм), средние (7-10 мкм) и большие лимфоциты (10-18 мкм).
(2). Функциональная классификация. К лимфоцитам относят сходные морфологически, но различающиеся функционально клетки. Выделяют следующие типы: В-лимфоциты, Т лимфоциты и NK-клетки. Важное практическое значение имеет также подразделение лимфоцитов по дифференцировочным Аг.
Продолжительность жизни. В-клетки образуются в костном мозге, Т-клетки созревают в вилочковой железе. Продолжительность жизни лимфоцитов достаточно велика: от нескольких месяцев до нескольких лет.
Морфология: Малый лимфоцит имеет округлое с небольшими выемками ядро, содержащее конденсированный хроматин. Цитоплазма образует узкое кольцо вокруг ядра. Т-и В-лимфоциты морфологически относят к малым (большинство лимфоцитов в кровотоке).
Большой лимфоцит. Такие клетки составляют 3% общего количества циркулирующих в крови лимфоцитов. Ядро крупное и относительно плотное. В цитоплазме немного лизосом, небольшое количество митохондрий, рудиментарный комплекс Гольджи, минимум эндоплазматической сети и сравнительно большое количество свободных рибосом. Клетка образует короткие отростки /микроворсинки/,более многочисленные у В-лимфоцитов.
Большие, а также средние лимфоциты крови- активированные Аг В-лимфоциты, дифференцирующиеся в плазматические клетки,к большим лимфоцитам относят NK-клетки.
В-лимфоциты составляют менее 10% лимфоцитов крови. Эти клетки, а точнее клоныплазматических клеток способны вырабатывать против конкретных Аг соответстсвующие АТ. Каждый клон, происходящий из В-лимфоцитов, синтезирует и секретирует АТ только к одному Аг.
●. Цитотоксические Т-лимфоциты Тс, — , уничтожают инфицированные вирусом и чужеродные клетки при помощи перфорина. Взаимодействуют с молекулой ГКГ I класса в плазматической мембране клетки-мишени.
● Т-супрессоры (ТS) — регулируют интенсивность иммунного ответа, подавляя активность Тн клеток. Предотвращают развитие аутоиммунных реакций. Защищают организм от нежелательных последствий иммунной реакции, от чрезмерного воспаления и аутоагрессии. Т-супрессоры обеспечивают толерантность (невосприимчивость) матери к отцовским Аг, представленным на клетках плода, что даёт возможность выживать чужеродному в иммунологическом отношении плоду в организме матери.
● В- лимфоциты ответственны за гуморальный иммунный ответ. В мембране В-лимфоцита присутствует мономер IgМ. Из красного костного мозга В-лимфоциты мигрируют в тимус -независимые зоны лимфоидных органов. Продолжительность жизни большинства В-лимфоцитов не превышает десяти дней, если они не активируются Аг. Зрелые В-лимфоциты (плазматические клетки) вырабатывают АТ — Ig всех известных классов.
● NК-клетки не имеют поверхностных детерминант, характерных для Т- и В-лимфоцитов (ГКГ-нерестригированные киллеры). NК- клетки убивают ауто-, алло- , и ксеногенные опухолевые клетки, некоторые инфицированные вирусом и бактериями клетки. В типичных NК -клетках экспрессируются дифференцировочные Аг.
Нормальные киллеры, или клетки - убийиы большие лимфоциты с относительно рыхлым почковидным крупным ядром и обильной цитоплазмой в которой содержатся митохондрии, свободные рибосомы с элементами шероховатого эндоплазматического ретикулума.
ранулы с большим количеством цитотоксических веществ. На внешней мембране нормальных киллеров имеются специфические рецепторы, благодаря которым они, например, узнают злокачественные и инфицированные вирусом клетки и затем активируются. При этом разрушение клеток-мишеней, по-видимому, вызывает выделяемый ими перфорин, молекулы которого имеют такое же трубчатое строение, как комплемент или пептидные антибиотики бактерий валиомицин и грамицидин.
● Цитолиз. В отличие от цитотоксических Т-лимфоцитов, способность NК -клеток к цитолизу не связана с необходимостью распознавания молекул ГКГ на поверхности мишени. NК- клетки уничтожают клетку-мишень не путём фагоцитоза, а (после установления с ней прямого контакта) при помощи перфорина.
● Гуморальная регуляция. Активность NК-клеток регулируется цитокинами. γ-ИФН и ИЛ-2 усиливают цитолитическую активность NК-клеток.
● Участие в антитело -зависимом клеточно-опосредованном цитолизе. NК -клетки, наряду с макрофагами, нейтрофилами и эозинофилами, участвуют в АТ- зависимом клеточно-опосредованном цитолизе. Для этого NК -клетки экспрессируют на своей поверхности рецептор Fс- фрагмента IgG. Реакция зависит от присутствия АТ (Ig), узнающих клетку-мишень и связывающихся с ней. Fс -фрагмент связанных с клеткой-мишенью АТ взаимодействует с рецептором Fс- фрагмента, встроенным в плазматическую мембрану NК - клетки. Природа агента, убивающего клетку-мишень в этом случае, неизвестна.
● Аг- представляющие клетки: макрофаги, В-лимфоциты, фолликулярные отростчатые клетки лимфоузлов и селезёнки, клетки Лангерханса, М-клетки в лимфатических фолликулах пищеварительного тракта, дендритные эпителиальные клетки вилочковой железы.
Эти клетки захватывают, процессируют и представляют Аг (эпитоп) на своей поверхности другим иммунокомпетентным клеткам, вырабатывают ИЛ-1 и другие цитокины, секретируют простагландин, угнетающий иммунный ответ, усиливюет фагоцитарную и цитолитическую активность макрофагов.
ЛИМФО МИЕЛОИДНЫЙ КОМПЛЕКС.
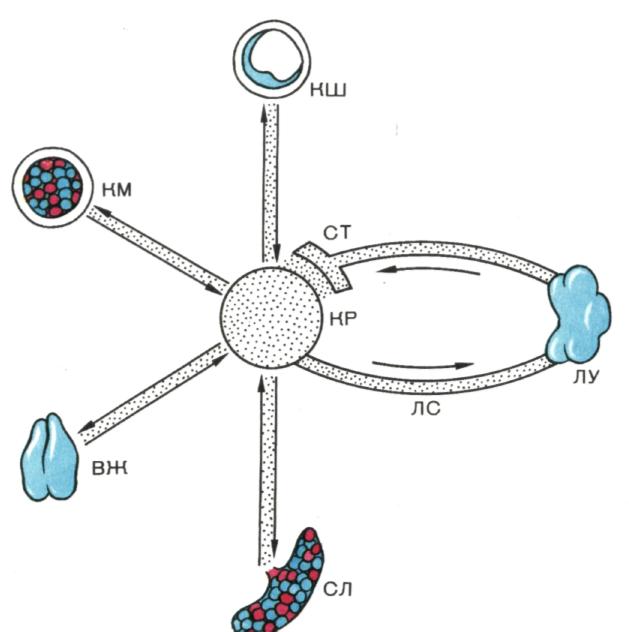
рис. 3. .Лимфо - миелоидныи комплекс.
Комплекс представляет собой систему органов и тканей, паренхима которых содержит клетки мезенхимального происхождения. Органы и ткани комплекса объединены в единую систему сетью кровеносных (КР) и лимфатических сосудов (ЛС). Комплекс включает: костный мозг (КМ), вилочковую железу (ВЖ), лимфатические узлы (ЛУ), лимфоидную ткань кишечника (КШ), селезенку (СЛ) и соединительную ткань (СТ). Функциональное назначение комплекса — обеспечение кроветворения (миелопоэза) и формирование клеток иммунной системы (лимфопоэза). Среди органов и тканей комплекса имеются истинно лимфоидные образования, в которых происходит только лимфопоэз (вилочковая железа, лимфатические узлы, лимфоидная ткань кишечника), и «смешанные» образования, в которых представлен как лимфо-, так и миелопоэз (костный мозг, селезенка).

Рис. 4. Кандидат на стволовую кроветворную клетку.
Стволовая кроветворная клетка дает начало всем росткам дифференцировки клеток лимфоидной и миелоидной ткани и тем самым формирует клеточный состав лимфомиелоидного комплекса. Морфология стволовой кроветворной клетки неизвестна. Однако при фракционировании клеток костного мозга в градиенте плотности альбумина и последующей оценке колониеобразующей способности клеток различных фракций представили возможного кандидата на стволовую кроветворную клетку. Эта клетка содержит округлое ядро (Я) с неглубокими вырезами, имеет два больших четко обозначенных ядрышка (Яд). Хроматин хлопкообразный или тонко дисперсный. Вдоль ядерной оболочки видны незначительные агрегаты гранул хроматина. Относительно тонкий ободок цитоплазмы содержит преимущественно свободные рибосомы. Имеется несколько цитоплазматических вакуолей (В). Митохондрии (М) относительно редки и невелики по размерам (X17 000).
.
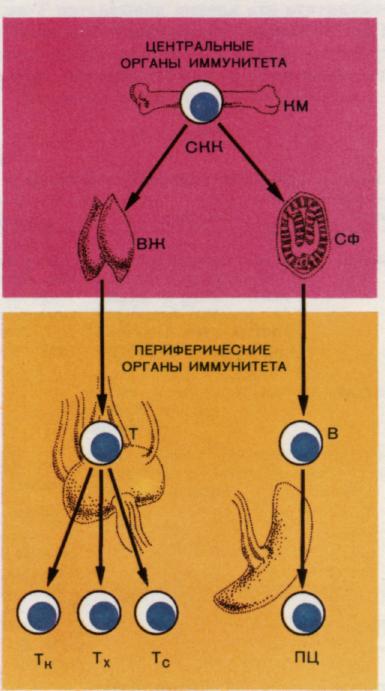
Рис. 5. Формирование Т- и В-систем иммунитета.
Стволовая кроветворная клетка (СКК), или клетка-предшественник лимфопоэза, мигрируя из костного мозга (КМ) в вилочковую железу (ВЖ) дифференцируется под влиянием микроокружения этого органа в Т-клетку. При миграции СКК в сумку Фабрициуса (СФ) у птиц или неизвестный аналог данного органа у млекопитающих происходит дифференцировка в клетки В-ряда. Из центральных, или первичных, органов иммунитета (костный мозг, вилочковая железа) клетки расселяются в периферические, или вторичные, органы иммунной системы (селезенку, лимфатические узлы и др.), где они приобретают функциональные особенности, свойственные зрелым Т- и В- клеточным популяциям. Т- киллер – клетка с цитолитической активностью по отношению к антигенно неродственной клетке- мишени. Т- хелпер – клетка, обеспечивающая положительную / усиливающую/ регуляцию гуморального иммунного ответа, в случае положительной регуляции клеточного иммунитета эта клетка называется Т- амплификатором. Т - супрессор – клетка, обеспечивающая негативную /подавляющую/ регуляцию иммунного ответа. ПЦ – плазмоцит, конечная клетка в В - клеточном пути дифференцировки, активно синтезирующая и секретирующая антитела.
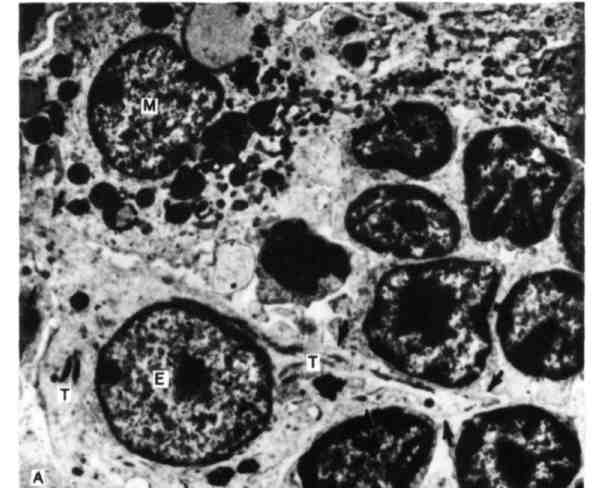
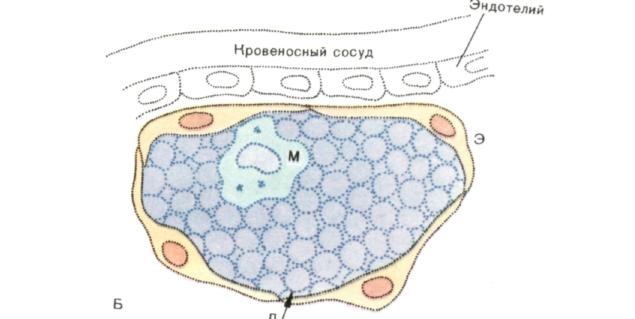
Рис. 6. Фолликул Кларка в вилочковой железе.
А. Элементарными структурно-гистологическими единицами вилочковой железы являются фолликулы Кларка. Они расположены главным образом в корковом веществе и включают эпителиальные клетки (Э), лимфоциты (Л) и макрофаги (М). Отличительными субклеточными структурами эпителии-альных клеток являются тонофибриллы (Т). Отростки эпителиальных клеток и псевдоподии макрофагов (отмечено стрелками) обеспечивают контакт этих клеток с лимфоцитами, входящими в состав фолликула.
Б. Реконструкция фолликула Кларка.
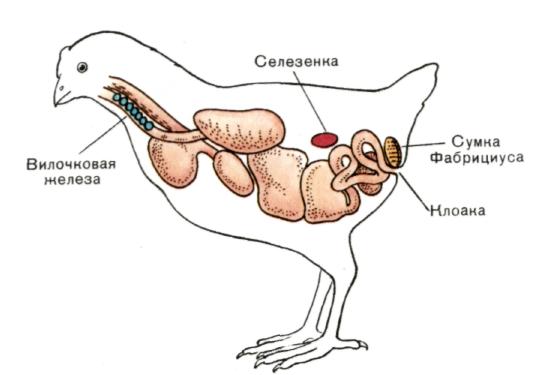
Рис. 7. Локализация сумки Фабрициуса у птиц.
Сумка Фабрициуса — лимфоэпителиальный орган, расположенный в задней части клоаки у птиц. В этом органе происходит генерация В-клеток. Аналог сумки Фабрициуса у млекопитающих пока не известен.

Рис. 8. Гистологическое строение сумки Фабрициуса.
Просвет сумки выстлан цилиндрическим эпителием, подобным эпителию кишечника. Неспосредственно за эпителиальным слоем располагаются узелки (дольки), общее строение которых неотличимо от строения соответствующих структур вилочковой железы. Кора представлена в основном плотным скоплением малых лимфоцитов. Более светлое мозговое вещество включает большие лимфоциты, плазматические клетки, макрофаги, гранулоциты, ретикулярные клетки. Эпителиальные клетки образуют сеть, переходящую
в эпителиальные покровы просвета органа. В отличие от вилочковой железы в узелках сумки корковый слой отделен от медуллярного основной мембраной. А — схематическое изображение участка сумки Фабрициуса;
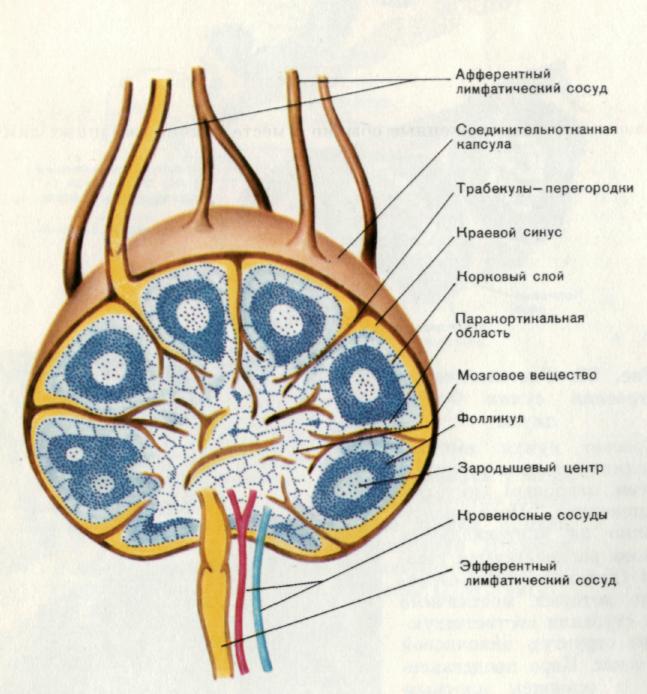
Рис. 9. Лимфатический узел.
Лимфатические узлы представляют собой хорошо различимые образования, расположенные обычно в месте слияния крупных лимфатических сосудов. Размеры узлов у человека в условиях нормы колеблются от 3 до 30 мм. Снаружи узел покрыт соединительнотканной капсулой. От капсулы в глубь узла отходят перегородки — трабекулы. Непосредственно под капсулой находится краевой синус, куда поступает лимфа из афферентных (приносящих) лимфатических сосудов. Из краевого синуса лимфа поступает в промежуточные синусы, пронизывающие всю толщу органа, и собирается в эфферентный (выносящий) лимфатический сосуд. Место выхода сосуда называется воротами узла. Через ворота в узел проходят кровеносные сосуды. Лимфоидная ткань узла делится на корковый слой и мозговое вещество. Корковый слой характеризуется плотной упаковкой лимфоидных клеток, которые собраны в округлые скопления — первичные и вторичные фолликулы. Первичные фолликулы представляют собой естественные гистологические структуры органа. Вторичные фолликулы отличаются наличием светлой центральной части, состоящей из пролиферирующих бластных форм. Они получили название зародышевых центров и образуются в ответ на проникновение в орган антигена.
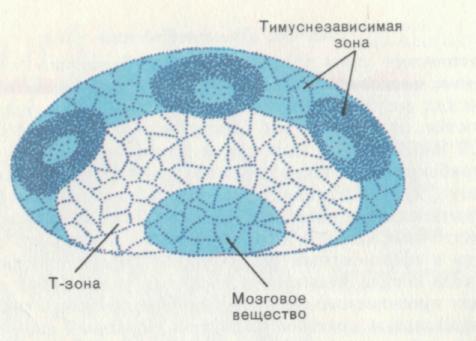
Рис. 10. Тимусзависимые и тимуснезависимые зоны в лимфатическом узле.
Неонатальная тимэктомия у мышей приводит к исчезновению лимфоцитов из паракортикальной области, которая получила название тимусзависимой зоны (Т-зоны). Кора и мозговое вещество при тимэктомии остаются незатронутыми. Сделан вывод, что лимфоциты вилочковой железы заселяют Т-зону, а лимфоциты костного мозга — В-зону, или тимуснезависимую зону.
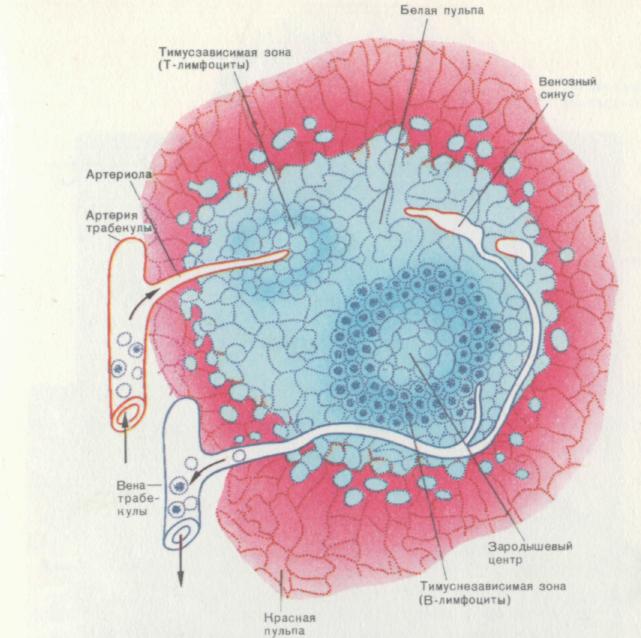
рис. 11. Участок селезенки (схема).
Селезенка относится к периферическим органам иммунной системы. Снаружи орган окружен соединительнотканной капсулой, от которой отходят поддерживающие перегородки — трабекулы. Паренхима селезенки включает белую и красную пульпы. Белая пульпа — место локализации лимфоцитов, которые собраны в отдельные элементарные гистологические структуры (мальпигиевы тельца). Красная пульпа состоит из ретикулокапиллярных петель, пространство между которыми заполнено свободными клеточными элементами. Среди этих клеток большинство представлено эритроцитами, что и определяет цвет пульпы. Четких границ между белой и красной пульпой нет, и переход клеточного состава из одной области в другую происходит постепенно. Белая пульпа заселяется Т- и В-лимфоцитами, мигрирующими из центральных органов иммунной системы. Они распределяются по двум зонам: тимусзависимой, где скапливаются Т-лимфоциты вокруг пронизывающих пульпу артериол, и тимуснезависимой — места накопления В-Лимфоцитов. В этой зоне хорошо различимы фолликулы с центрами размножения, которые образуются в ответ на антигенный стимул.
Рис. 12. Этапы антигеннезависимой дифференцировки Т-клеток.
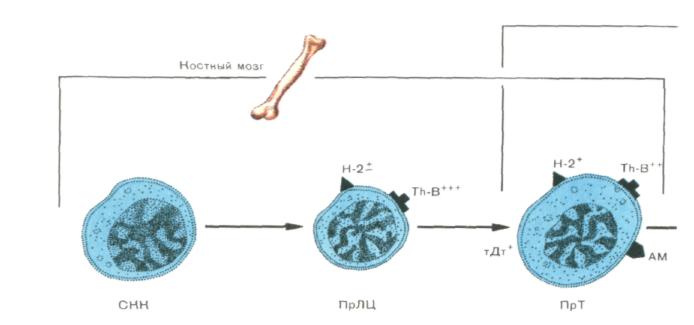
На первом этапе из полипотентной стволовой кроветворной клетки (СКК) костного мозга, обеспечивающей миело- и лимфопоэз, образуется общий для Т- и В-лимфоцитов предшественник (ПрЛЦ). Такой коммитированный в сторону лимфопоэза предшественник становится обладателем антигенов Н-2 и антигена Тh-В. Предполагается, что этот антиген является стадиоспеци-фическим маркером ПрЛЦ. Его количество снижается по мере созревания лимфоцитов, вышедших на Т-клеточный путь развития. Ближайшим потомком ПрЛЦ является протимоцит. Маркером данного клеточного типа служит терминальная дезокеинуклеотидилтрансфераза (тДт), выявленная только у кортикальных тимоцитов. Другой отличительный маркер ПрТ — один из антигенов мозга (Ам). ПрТ локализуются в костном мозге, а также непосредственно под капсулой вилочковой железы, где их содержание приближается к 5%.
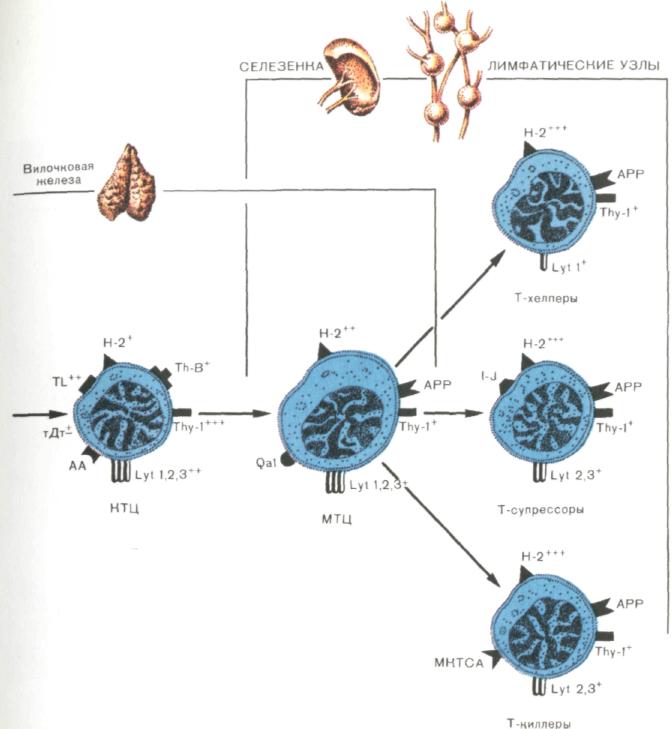
Под влиянием гормонов вилочковой железы, секретируемых эпителиальными клетками, и монокинов, выделяемых тимоцитарными макрофагами, происходит созревание ПрТ до кортикальных тимоцитов (КТЦ). КТЦ становятся обладателями хорошо выраженного специфического антигена Т-клеток Тhу-1, а также дифференцировочных антигенов Lу1,1, 2, 3. Активность тДт снижается по сравнению с таковой в клетках предшествующей стадии дифференцировки (ПрТ).. Прямые потомки кортикальных клеток — медуллярные тимоциты (МТЦ), располагающиеся в мозговом веществе вилочковой железы и мигрирующие в периферические лимфоидные органы. МТЦ характеризуются большими размерами по сравнению с КТЦ, лучшей выраженностью антигенов Н-2, отсутствием маркера.. Ранние этапы дифференцировки в костном мозге и направляющей по Т-клеточному пути развития дифференцировки в вилочковой железе завершаются на периферии (селезенка, лимфатические узлы) образованием функционально зрелых субпопуляций Т-клеток: Т-киллеров, Т-супрессоров и Т-хелперов.
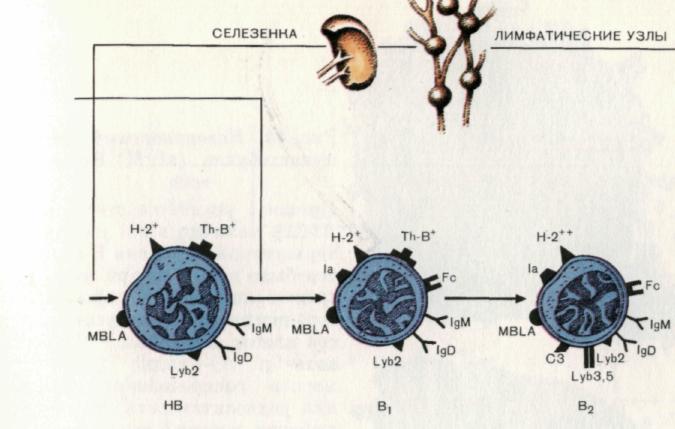
Рис. 13. Этапы антигеннезависимой дифференцировки В-клеток.
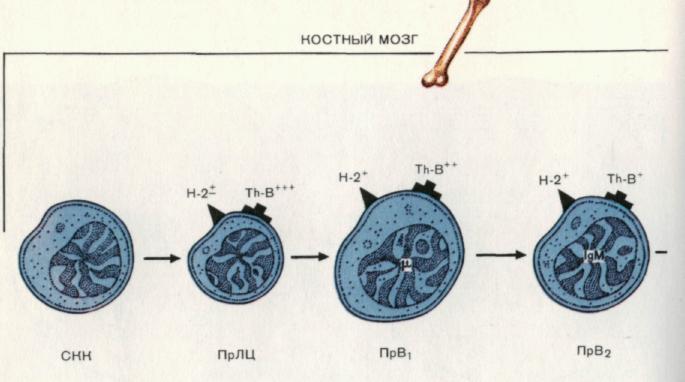
На первых этапах дифференцировки на территории костного мозга взрослых мышей от общего прекурсора для Т- и В-лимфоцитов (ПрЛЦ) формируется родоначальная форма для В-клеточного пути развития, или предшественник В-клеток первого типа (ПрВ1). В данном клеточном типе происходит активация гена для тяжелой цепи IgМ. Продукт этого гена представлен только в цитоплазме и отсутствует на клеточной поверхности. Количество Тh-В-антигена снижено по сравнению с количеством на ПрЛЦ.
На следующем этапе раннего периода развития образуется предшественник В-клеток 2-го типа (ПрВ2). Основным признаком клеток этого типа является наличие в цитоплазме мономерных молекул IgМ и их отсутствие на клеточной поверхности. Уровень Тh-В-антигена прогрессивно снижается. Первые наиболее ранние этапы дифференцировки генерализуют процесс развития по В- клеточному пути.
Незрелая В-клетка (НВ) представлена как в костном мозге, так и на периферии, главным образом в селезенке, хотя небольшое их количество имеется и в лимфатических узлах. Для данного этапа дифференцировки характерны появление маркера В-клеток — МВLА, наличие поверхностных иммуноглобулинов двух классов — IgМ и IgD, отсутствие маркеров зрелых В-клеток. НВ способны к распознаванию антигена. Однако их контакт с антигенами не приводит к трансформации в антителопродуценты.
В2-клетки являются непосредственными предшественниками антителопро-дуцирующих клеток, формирующихся после контакта В2-клеток с антигеном.
Рис. 14. Этапы созревания тканевого макрофага от прекурсора костного мозга.
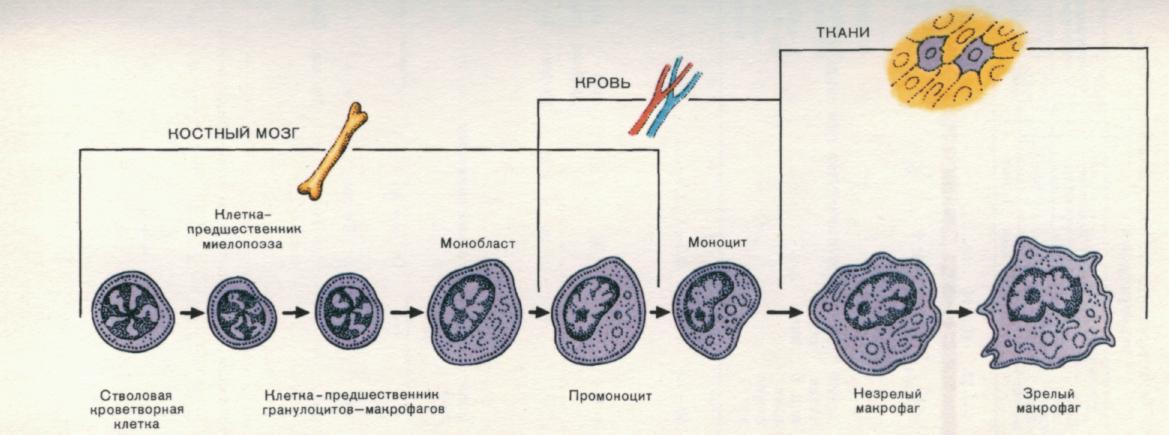
Первые этапы дифференцировки происходят в костном мозге, где определяется линия развития в направлении макрофагального ростка. В крови основной представитель данной линии развития — моноцит. При его проникновении в паренхиму органов осуществляются завершающие этапы дифференцировки, которые приводят к формированию зрелого не вступающего в пролиферацию тканевого макрофага.
Дата добавления: 2017-01-16; просмотров: 3092;
Поиск по сайту
Узнать еще
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- I.2. Антигены системы АВ0. Генетика. Структура
- I.2.1 ПЕРВЫЙ ЗАКОН НЬЮТОНА. ИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА.
- I.2.1 ПОЛНАЯ И ВНУТРЕННЯЯ ЭНЕРГИЯ СИСТЕМЫ. ТЕПЛОТА И РАБОТА
- I.3. Антитела системы АВ0
- I.5.4 НЕИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА. СИЛЫ ИНЕРЦИИ
- II. ЭЛЕКТРИЧЕСКИЙ ДИПОЛЬ. ДИПОЛЬНЫЙ МОМЕНТ СИСТЕМЫ ЭЛЕКТРИЧЕСКИХ ЗАРЯДОВ
- III. Избирательные системы.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
