Особенности пыльцевой трубки
Двойное оплодотворение цветковых растений способствует прорастанию пыльцы. Оно начинается с разбухания зерна и последующего формирования пыльцевой трубки. Она прорывает спородерму в наиболее тонком ее участке. Называется он апертура. С кончика пыльцевой трубки выделяются специфические вещества. Они размягчают ткани столбика и рыльца. За счет этого в них входит пыльцевая трубка. По мере ее развития и роста, в нее переходят оба спермия и ядро от вегетативной клетки. В подавляющем большинстве случаев проникновение пыльцевой трубки в нуцеллус (мегаспорангий) происходит посредством микропиле семязачатка. Крайне редко это осуществляется другим способом. После проникновения в зародышевый мешок происходит разрыв пыльцевой трубки. В результате все ее содержимое изливается вовнутрь. Двойное оплодотворение цветковых растений продолжается формированием диплоидной зиготы. Этому способствует первый спермий. Второй элемент соединяется с вторичным ядром, которое расположено в центральной части зародышевого мешка. Образованное триплоидное ядро впоследствии трансформируется в эндосперм.
Двойное оплодотворение сопровождается взаимодействием ее с тканями спорофита. Оно достаточно специфично. Этот процесс регулируется активностью химических соединений. Установлено, что если пыльцу промыть в дистиллированной воде, она потеряет способность к прорастанию. Если же полученный раствор сконцентрировать, а затем ее обработать, то она снова станет полноценной. Развитие пыльцевой трубки после прорастания контролируют ткани пестика. К примеру, у хлопчатника ее рост до яйцеклетки занимает порядка 12-18-ти часов. Однако уже спустя 6 часов вполне можно определить, к какой именно семяпочке будет направляться пыльцевая трубка. Это понятно потому, что в ней начинается разрушение синергиды. В настоящее время не установлено, как растение может направить развитие трубки в нужном направлении и каким образом о приближении узнает синергида.
"Запрет" на самоопыление
Он достаточно часто наблюдается у цветковых растений. Это явление имеет свои особенности. "Запрет" на самоопыление проявляется в том, что спорофит "идентифицирует" собственного мужского гематофита и не допускает его к участию в оплодотворении. При этом в ряде случаев на рыльце пестика не происходит прорастания собственной пыльцы. Однако, как правило, рост трубки все-таки начинается, но впоследствии приостанавливается. В результате пыльца не достигает яйцеклетки и, как следствие, двойное оплодотворение не происходит. Еще Дарвиным было отмечено это явление. Так, он обнаружил у первоцвета весеннего цветки двух форм. Одни из них были длинностолбиковыми с короткими тычинками. Другие же – короткостолбиковыми. В них тычиночные нити были длинные. Короткостолбиковые растения отличаются крупной пыльцой (вдвое больше, чем у других). При этом клетки в сосочках рыльца – мелкие. Указанные признаки контролирует группа из тесно сплетенных генов.
Рецепторы
Двойное оплодотворение эффективно, когда пыльца переносится от одной формы к другой. За распознавание собственных элементов отвечают особые молекулы-рецепторы. Они представляют собой сложные соединения углеводов с белками. Установлено, что формы дикой капусты, не вырабатывающие в тканях рыльца эти молекулы-рецепторы, способны самоопыляться. Для нормальных растений характерно появление углеводно-белковых соединений за день до раскрытия цветка. Если открыть бутон и обработать его собственной пыльцой за двое суток до его распускания, то двойное оплодотворение произойдет. Если это сделать за день до открытия, то его не будет.
Аллели
Примечательно, что в ряде случаев "самонесовместимость" пыльцы в растениях устанавливается серией множественных элементов одного гена. Это явление похоже на несовместимость при пересадке ткани у животных. Такие аллели обозначают литерой S. Число в популяции этих элементов может достичь десятков или даже сотен. К примеру, если генотип растения, производящего яйцеклетки, - s1s2, а вырабатывающего пыльцу - s2s3, при перекрестном опылении прорастание будет отмечено только у 50% пылинок. Это будут те, которые несут аллель s3. Если элементов несколько десятков, то большая часть пыльцы прорастет нормально при перекрестном опылении, при этом самоопыление предотвращается полностью.
В заключение
В отличие от голосеменных, для которых характерно развитие достаточно мощного гаплоидного эндосперма вне зависимости от оплодотворения, у покрытосеменных ткань образуется только в этом единственном случае. Учитывая огромное количество поколений, таким образом достигается значительная экономия энергии. Повышение степени плоидности эндосперма, по всей видимости, способствует более скорому росту ткани в сравнении с диплоидными слоями спорофита.
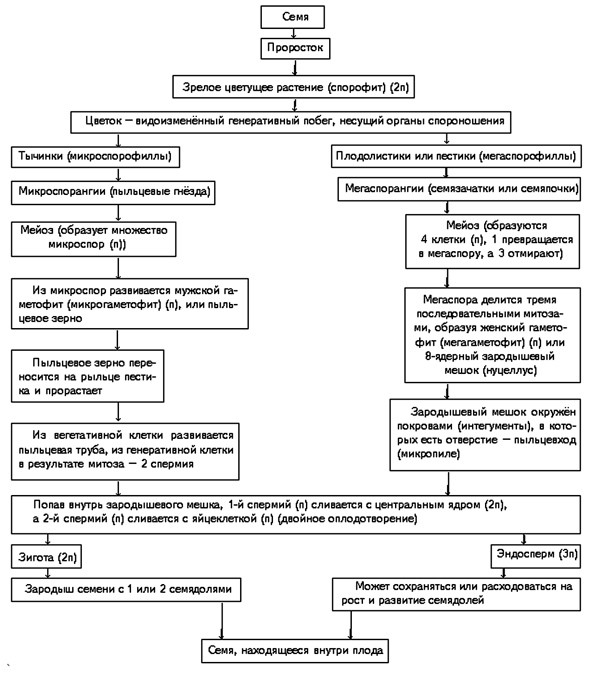
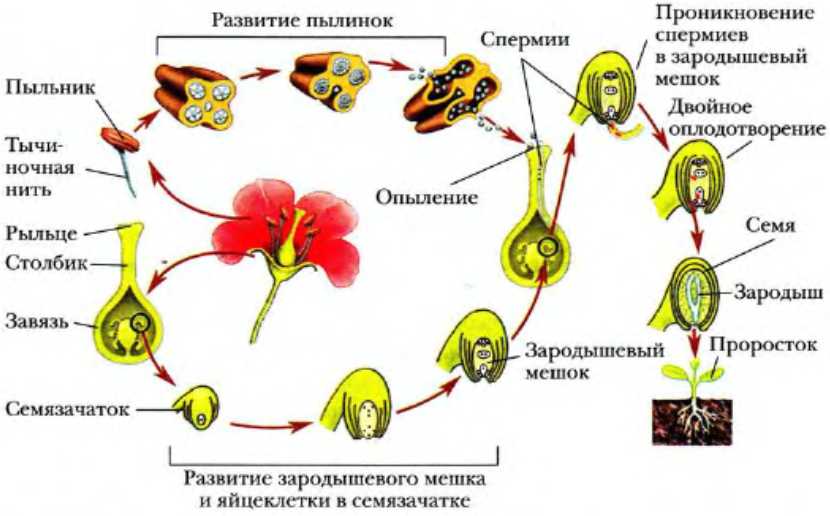
Строение семени.
Дата добавления: 2017-01-08; просмотров: 3774;
Поиск по сайту
Узнать еще
- I. ОСОБЕННОСТИ ДЕЛОВОГО И ЛИЧНОСТНОГО ОБЩЕНИЯ В СОВМЕСТНОЙ ДЕЯТЕЛЬНОСТИ
- I.2. Основные категории водопотребления промышленных предприятий и их особенности
- I2. Особенности аэродинамики несущего винта (НВ)
- II. Завоевание Китая маньчжурами. Экономическое положение страны в XVII – начале XIX вв.: аграрная политика Цинской династии, особенности развития городского ремесла
- II. Особенности политического устройства Ирана
- II. Особенности развития турецкой буржуазии. Становление младотурецкого движения
- II. РЕЖИМ И ОСОБЕННОСТИ ЛИЧНОЙ ГИГИЕНЫ
- III. Особенности политической элиты России.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
