Биосинтез и метаболизм кортикостероидов
Общим предшественником кортикостероидов служит холестерол (рис. 11-20).
В митохондриях холестерол превращается в прегненолон при участии гидроксилазы, относящейся к группе цитохромов Р450. Цитохром Р450, отщепляющий боковую цепь, локализован во внутренней мембране митохондрий. Отщепление боковой цепи холестерола включает 2 реакции гидроксилирования: одна - по атому С22, другая - по С20. Последующее отщепление шестиуглеродного фрагмента приводит к образованию С21-стероида - прегненолона. Дальнейшее превращение прегненолона происходит под действием различных гидроксилаз с участием молекулярного кислорода и NADPH, а также дегидрогеназ, изомераз и лиаз. Эти ферменты имеют различную внутри- и межклеточную локализацию. В коре надпочечников различают 3 типа клеток, образующих 3 слоя, или зоны: клубочковую, пучковую и сетчатую. Каким именно стероидом окажется конечный продукт, зависит от набора ферментов в клетке и последовательности реакций гидроксилиро-вания. Например, ферменты, необходимые для синтеза альдостерона, присутствуют только в клетках клубочковой зоны, а ферменты синтеза глюкокортикоидов и андрогенов локализованы в пучковой и сетчатой зонах.
Путь биосинтеза кортизола. Кортизол синтезируется из холестерола, который в основном поступает из крови в составе ЛПНП или синтезируется в клетках из ацетил-КоА. Значительная часть эфиров холестерола накапливается в цитозоле клеток в липидных каплях. Под влиянием АКТГ происходит активация специфической эстеразы,
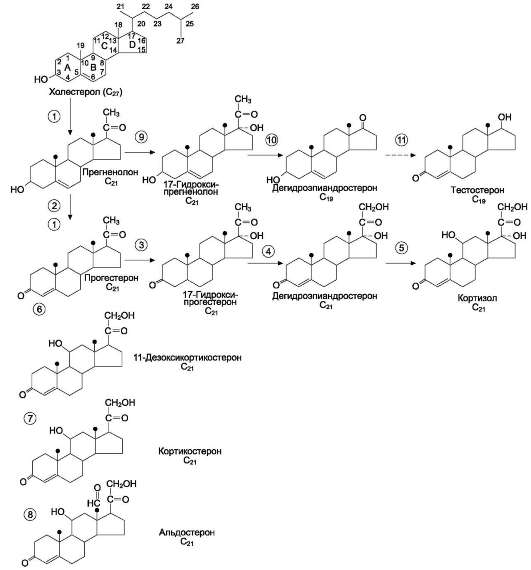
Рис. 11-20. Строение и основные этапы синтеза кортикостероидов.1 - превращение холестерола в прегненолон (гидроксилаза, отщепляющая боковую цепь); 2 - образование прогестерона (3-Ь-гидроксис-тероиддегидрогеназа); 3, 4, 5 - реакции синтеза кортизола (3 - 17-гидроксилаза, 4 - 21-гидроксилаза, 5 - 11-гидроксилаза); 6, 7, 8 - путь синтеза альдостерона
(6 - 21-гидроксилаза, 7 - 11-гидроксилаза, 8 - 18-гидроксилаза, 18-гидроксидегидрогеназа); 9, 10, 11 - путь синтеза тестостерона (9 - 17-гидроксилаза, 10 - 17,20-лиаза, 11 - дегидрогеназа).
и свободный холестерол транспортируется в митохондрии (рис. 11-21).
Синтез кортизола начинается с превращения прегненолона в прогестерон. Эта реакция протекает в цитозоле клеток пучковой зоны коры надпочечников, куда прегненолон транспортируется из митохондрий. Реакцию катализирует 3-β-гидроксистероиддегидрогеназа.
В мембранах ЭР при участии 17-α-гидрок-силазы происходит гидроксилирование про-гес-терона по С17 с образованием 17-гидрокси-прогестерона. Этот же фермент катализирует превращение прегненолона в 17-гидроксип-регненолон, от которого далее при участии
17,20-лиазы может отщепляться двухуглеродная боковая цепь с образованием С19-стероида - дегидроэпиандростерона. 17-гидроксипрогес-терон служит предшественником кортизола, а дегидроэпиандростерон - предшественником андрогенов. Далее 17-ОН-прогестерон гидрок-силируется 21-гидроксилазой (Р450-С21), локализованной в мембране ЭР, и превращается в 11-дезоксикортизол, который переносится во внутреннюю мембрану митохондрий, где гид-роксилируется при участии цитохрома Р450-С11 с образованием кортизола.
Скорость синтеза и секреции кортизола стимулируются в ответ на стресс, травму, ин-
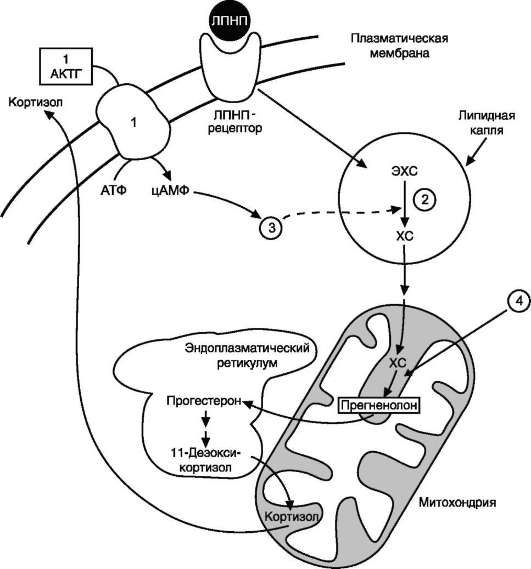
Рис. 11-21. Внутриклеточная локализация синтеза кортизола.1 - аденилатциклазный комплекс; 2 - хо-лестеролэстераза; 3 - протеинкиназа А; 4 - холестеролдесмолаза отщепляет боковую цепь холестерола. ХС - холестерол; ЭХС - эфиры холестерола.
фекцию, понижение концентрации глюкозы в крови. Повышение концентрации кортизола подавляет синтез кортиколиберина и АКТГ по механизму отрицательной обратной связи.
Синтез минералокортикоидов в клетках клу-бочковой зоны коры надпочечников также начинается с превращения холестерола в пре-гненолон, а затем в прогестерон. Прогестерон гидроксилируется вначале по С21 с образованием 11-дезоксикортикостерона. Следующее гидроксилирование происходит по С11, что приводит к образованию кортикостерона, обладающего слабовыраженной глюкокортикоидной и минералокортикоидной активностью.
В клетках клубочковой зоны 17-а-гидроксилаза отсутствует, но есть митохондриальная 18-гид-роксилаза, при участии которой кортикостерон гидроксилируется, а затем дегидрируется с образованием альдегидной группы у С18.
Главным стимулом для синтеза альдостерона служит ангиотензин II (см. ниже подраздел V).
Транспорт кортикостероидов. Кортизол в плазме крови находится в комплексе с α-глобулином транскортином и в небольшом количестве в свободной форме. Синтез транскортина протекает в печени и стимулируется эстрогенами.
T1/2 кортизола составляет 1,5-2 ч. Несвязанный, или свободный кортизол, составляет около 8% от общего количества гормона в плазме и является биологически активной фракцией.
Альдостерон не имеет специфического транспортного белка, но образует слабые связи с альбумином.
Катаболизм гормонов коры надпочечников происходит прежде всего в печени. Здесь протекают реакции гидроксилирования, окисления и восстановления гормонов. Продукты катаболизма кортикостероидов (кроме кортикостерона и альдостерона) выводятся с мочой в форме 17-кетостероидов, образующихся в результате отщепления боковой цепи. Эти продукты метаболизма выделяются преимущественно в виде конъюгатов с глюкуроновой и серной кислотами. 17-Окси- и 17-кетостероиды образуются также при катаболизме половых гормонов, которые имеют у С17гидроксиили кетогруппы. У мужчин 2/3 кетостероидов образуется за счёт кортикостероидов и 1/3 за счёт тестостерона (всего 12-17 мг/сут). У женщин 17-кетосте-роиды образуются преимущественно за счёт кортикостероидов (7-12 мг/сут). Определение
17-кетостероидов в моче позволяет оценить как количество глюкокортикоидов, секретируемых корой надпочечников, так и функцию надпочечников.
2. Биологические функции кортикостероидов отличаются широким спектром влияний на процессы метаболизма и подробно рассматриваются в соответствующих разделах.
Важнейший фактор в механизме действия кортикостероидов - взаимодействие их со специфическими рецепторами, расположенными в цитозоле клетки или в ядре. Регуляция внутриклеточных процессов под влиянием кортико-стероидных гормонов проявляется в изменении количества белков, обычно ключевых ферментов метаболизма, путём регуляции транскрипции генов в клетках-мишенях.
Влияние глюкокортикоидовна промежуточный метаболизм связано с их способностью коор-динированно воздействовать на разные ткани и разные процессы, как анаболические, так и катаболические.
Кортизол стимулирует образование глюкозы в печени, усиливая глюконеогенез и одновременно увеличивая скорость освобождения аминокислот - субстратов глюконеогенеза из периферических тканей. В печени кортизол индуцирует синтез ферментов катаболизма аминокислот (аланинаминотрансферазы, триптофанпирро-лазы и тирозинаминотрансферазы и ключевого фермента глюконеогенеза - фосфоенолпиру-ваткарбоксикиназы). Кроме того, кортизол стимулирует синтез гликогена в печени и тормозит потребление глюкозы периферическими тканями. Это действие кортизола проявляется в основном при голодании и недостаточности инсулина (см. ниже подраздел V). У здоровых людей эти эффекты кортизола уравновешиваются инсулином.
Избыточное количество кортизола стимулирует липолиз в конечностях и липогенез в других частях тела (лицо и туловище). Кроме того, глюкокортикоиды усиливают липолитическое действие катехоламинов и гормона роста.
Влияние глюкокортикоидов на обмен белков и нуклеиновых кислот проявляется двояко: в печени кортизол в основном оказывает анаболический эффект (стимулирует синтез белков и нуклеиновых кислот). В мышцах, лимфоид-ной и жировой ткани, коже и костях кортизол тормозит синтез белков, РНК и ДНК и стимулирует распад РНК и белков.
При высокой концентрации глюкокорти-коиды подавляют иммунные реакции, вызывая гибель лимфоцитов и инволюцию лимфоидной ткани; подавляют воспалительную реакцию, снижая число циркулирующих лейкоцитов, а также индуцируя синтез липокортинов, которые инги-бируют фосфолипазу А2, снижая таким образом синтез медиаторов воспаления - простагланди-нов и лейкотриенов (см. раздел 8).
Высокая концентрация глюкокортикоидов вызывает торможение роста и деления фибро-бластов, а также синтез коллагена и фибро-нектина (см. раздел 15). Для гиперсекреции глюкокортикоидов типичны истончение кожи, плохое заживление ран, мышечная слабость и атрофия мышц.
Глюкокортикоиды участвуют в физиологическом ответе на стресс, связанный с травмой, инфекцией или хирургическим вмешательством. В этом ответе в первую очередь участвуют кате-холамины, но во многих случаях для проявления их максимальной активности необходимо участие глюкокортикоидов.
Минералокортикоидыстимулируют реабсорб-цию Na+ в дистальных извитых канальцах и собирательных трубочках почек. Кроме того, они способствуют секреции К+, NH4+ в почках, а также в других эпителиальных тканях: потовых железах, слизистой оболочке кишечника и слюнных железах. В организме человека альдостерон - наиболее активный минералокортикоид.
Механизм действия и биологические эффекты альдостерона подробно рассмотрены в подразделе VI этого раздела.
Дата добавления: 2017-01-08; просмотров: 6253;
Поиск по сайту
Узнать еще
- III. Биосинтез крахмала и гликогена.
- Iii. строение, биосинтез и биологическое действие гормонов
- Активация аминокислот при биосинтезе белка.
- Анаболизм прокариот. Биосинтез углеводов
- Биосинтез аминокислот
- Биосинтез аминокислот
- Биосинтез белка в клетке
- Биосинтез белка и нуклеиновых кислот. Матричный характер реакций биосинтеза. Генетическая информация в клетке. Гены, генетический код и его свойства

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
