Класс Плазмодиофоромицеты
(Plasmodiophoromycetes)
Представители класса являются внутриклеточными паразитами. В отличие от свободноживущих слизевиков они не образуют мейоспорангиев, а мейоспоры развиваются из плазмодия внутри клетки растения-хозяина, которая выполняет функцию своеобразного вместилища спор. Плазмодиофоромицеты — паразиты, вызывающие гипертрофию тканей растения и образование опухолей.
Класс включает всего 1 порядок Плазмодиофоральные (Plasmodiophorales) и 4 рода, важнейшими из которых являются плазмодиофора и спонгоспора.
Род плазмодиофора (Plasmodiophora) включает виды с бесцветным зернистым плазмодием, распадающимся при созревании на отдельные, не объединенные в цистосорусы одноклеточные, шаровидные, мелкие споры с тонкой бесцветной, гладкой или шиповатой оболочкой. При прорастании споры обычно превращаются в зооспоры с длинным и коротким жгутиками. Род включает 6 видов, распространенных в умеренных поясах земного шара.
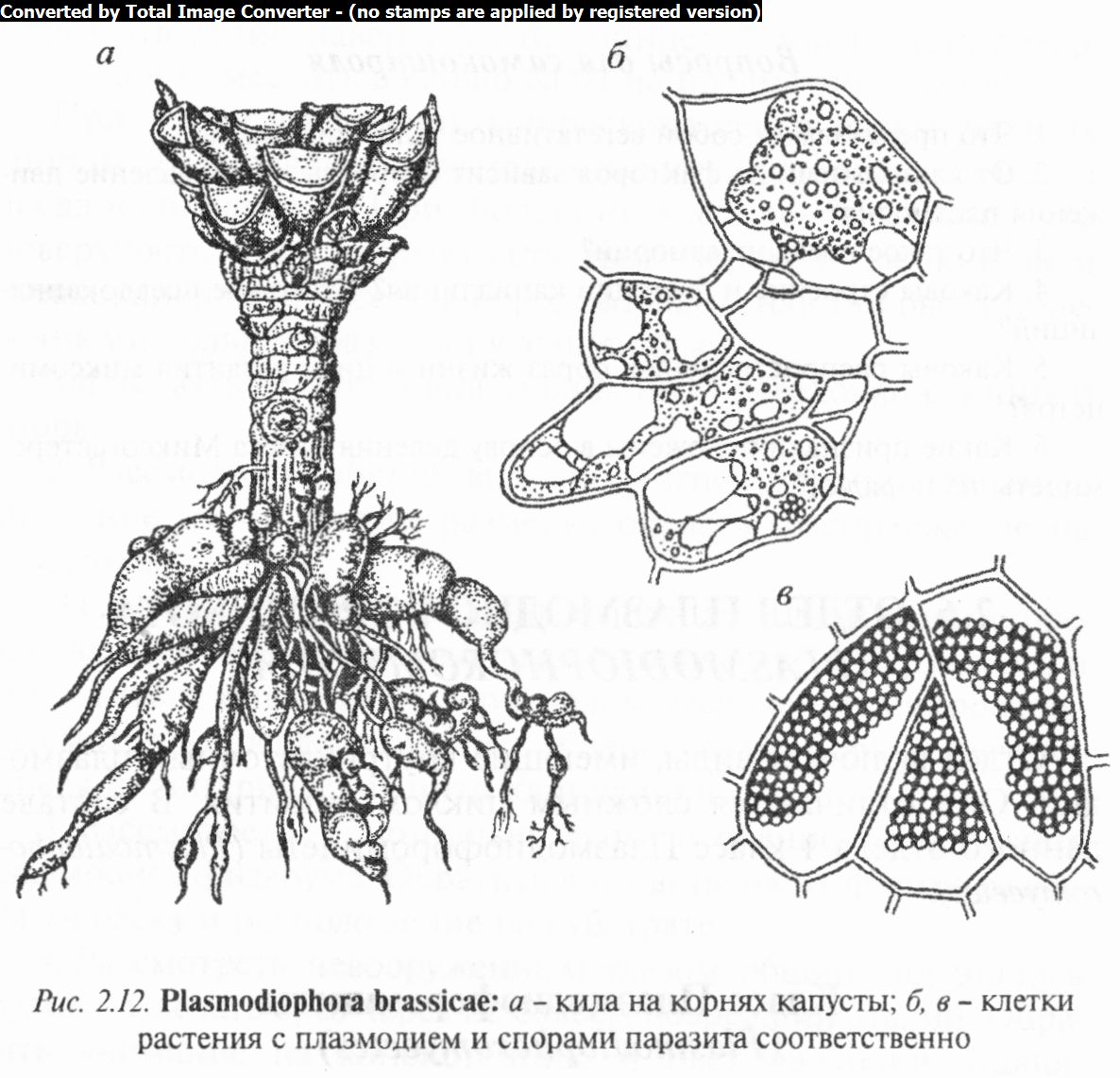
Наиболее известный представитель — П. капустная (P. brassicae) - возбудитель болезни корней капусты и других крестоцветных, так называемой килы. На корнях пораженного растения образуются наросты и вздутия, постепенно развивающиеся в очень крупные опухоли самой разнообразной формы (рис. 2.12). Такие корни почти не ветвятся и мало всасывают воды. У более взрослых растений листья становятся вялыми, желтоватыми, кочаны недоразвиваются или совсем не образуются.
Цикл развития возбудителя килы капусты протекает по следующей схеме. При сгнивании корней капусты споры слизевика попадают в почву, где они могут сохраняться годами. Для их прорастания нужны благоприятные условия влажности и температуры. Образовавшиеся при прорастании спор зооспоры или миксоамебы проникают в корневые волоски капусты, где происходит слияние миксоамеб (плазмогамия). В результате осуществляется формирование первичного многоядерного плазмодия, из которого развиваются зооспорангий (или гаметангии) и в них — зооспоры (или гаметы), выходящие из корне-
вого волоска в почву. Они попарно сливаются (без слияния ядер). Образующаяся при этом двухъядерная клетка заражает корень капусты и дает начало новым вторичным плазмодиям. В них многочисленные гаплоидные ядра, образующиеся в результате митотических делений, попарно сливаются, затем после мейотического деления диплоидных ядер плазмодии распадаются на многочисленные (до 25 ООО) покоящиеся споры, освобождающиеся после сгнивания корней.
Род спонгоспора (Spongospora) включает всего 1 вид — С. пасленовая (S. solani), поражающая клубни, корни, реже столоны растений картофеля, томатов и других пасленовых.
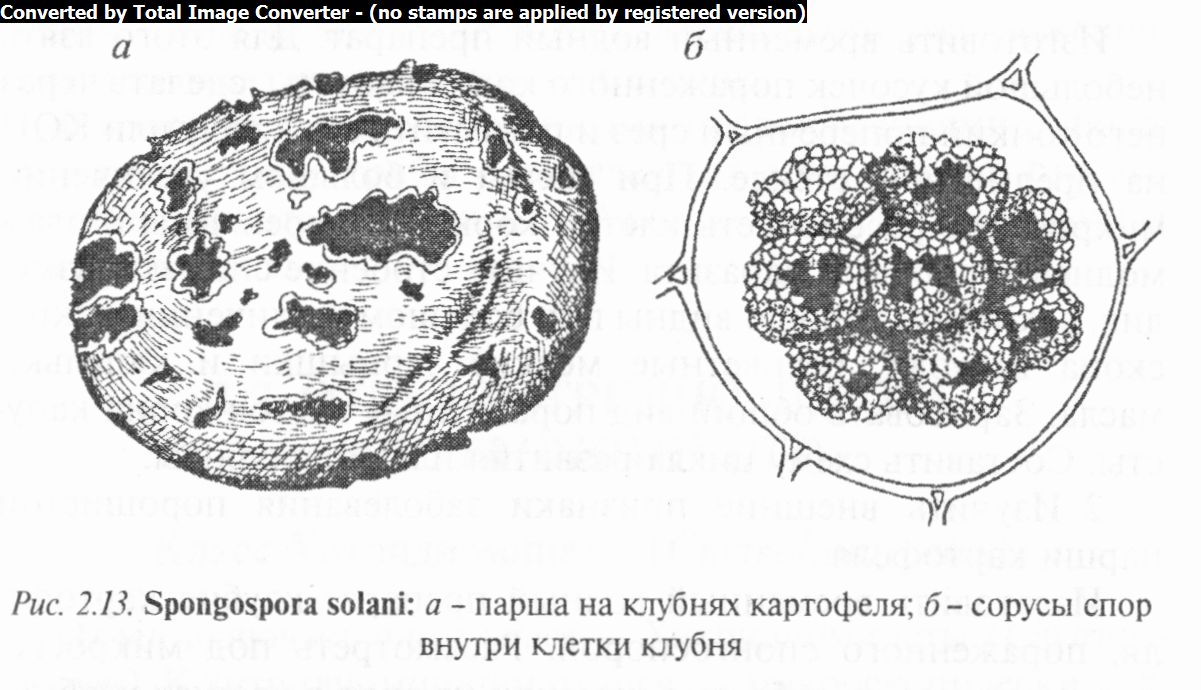
Спонгоспора — возбудитель порошистой парши картофеля. Многоядерный плазмодий в клетках покровных тканей клубня картофеля распадается на многочисленные овальные, бурые споры, собранные обычно в округлые сорусы. На поврежденных местах клубня сначала появляются бугорки, которые затем разрываются и обнажают цистосорусы с губчатыми комочками цист или спор (рис. 2.13). В почве покоящиеся споры прорастают зооспорами, которые проникают в растения через корневые волоски, чечевички, трещины в кожуре клубня и т.д. Заболевание известно почти во всех странах умеренного пояса, в том числе и в Беларуси.
Задания
1. Рассмотреть пораженные плазмодиофорой корни капусты на фиксированном материале.
Изготовить временный водный препарат. Для этого взять небольшой кусочек пораженного корня капусты, сделать через него тонкий поперечный срез и поместить его в воду или КОН на предметном стекле. При малом и большом увеличении микроскопа рассмотреть клетки корневой паренхимы с плазмодием и спорами паразита. Изучить строение спор и плазмодия, в котором хорошо видны при большом увеличении микроскопа вакуоли, бесцветные мелкие зернышки и капельки масла. Зарисовать общий вид пораженных клеток корня капусты. Составить схему цикла развития плазмодиофоры.
2. Изучить внешние признаки заболевания порошистой парши картофеля.
Изготовить временный водный препарат клубня картофеля, пораженного спонгоспорой. Рассмотреть под микроскопом и зарисовать губчатые комочки из спор в клетках клубня.
Вопросы и задания для самоконтроля
1. Охарактеризуйте циклы развития плазмодиофоры капустной, вызывающей килу, и спонгоспоры пасленовой — возбудителя порошистой парши картофеля.
2. Каково филогенетическое положение слизевиков?
3. Каково значение миксомицетов в природе и хозяйственной деятельности человека?
ЦАРСТВО НАСТОЯЩИЕ ГРИБЫ (FUNGI, MYCOTA, MYCETALIA)
Большинство грибов царства имеет настоящий мицелий. Каждая его гифа окружена тонкой жесткой стенкой, основным компонентом которой является хитин в сочетании с (3-глю- каном, маннаном или с хитозаном (в отделе Zygomycota). Для настоящих грибов характерно гетеротрофное абсорбтив- ное питание, неподвижность в вегетативном состоянии, наличие жгутиконосных клеток (спор и гамет) с одним (у представителей отдела Chytridiomycota) или несколькими жгутиками.
Классификация грибов основана на использовании комплекса признаков, из которых ведущими являются количество, строение и расположение жгутиков у грибов с подвижными стадиями развития, характер развития спор полового размно жения, типы полового процесса и бесполого размножения, состав полисахаридов клеточных стенок.
В соответствии с современными представлениями о филогении настоящие грибы распределены в следующие отделы: Хитридиомикота, Зигомикота, Аскомикота, Базидиомикота и Дейтеромикота.
2.7. ОТДЕЛ ХИТРИДИОМИКОТА (СН YTRIDIOM YCO ТА)
Класс Хитридиомицеты (Chytridiomycetes)
Отдел включает один класс - Хитридиомицеты (Chytridiomycetes). К хитридиомицетам относятся микроскопически мелкие грибы — самые простые по степени развития вегетативного тела, представленного голой плазменной массой, похожей на плазмодий слизевиков, или слаборазвитым мицелием (ризомицелием) в виде тонких, простых или ветвящихся безъядерных нитей, отходящих от основной округлой или удлиненной клетки таллома. Ризомицелий служит для прикрепления таллома к субстрату и поглощения из него питательных веществ. Виды с ризомицелием могут быть моно- или полицентрическими, т.е. иметь один или несколько центров формирования ризомицелия и репродуктивных органов (зооспорангиев и гаметангиев). В состав клеточной стенки хитридиомицетов входит комплекс — хитин с р-глюканом. Для некоторых хитридиомицетов характерен хорошо развитый несептированный мицелий с дифференцировкой его ветвей на ризоидальную, осевую и репродуктивные части.
Бесполое размножение осуществляется зооспорами различного строения и формы с одним бичевидным гладким жгутиком, прикрепленным сзади. У некоторых хитридиомицетов все вегетативное тело превращается в спорангий и жизнь данной особи прекращается (так называемые холокарпические формы), у других только часть таллома становится зооспоран- гием и таким образом зооспорангии могут развиваться по нескольку одновременно или последовательно (эукарпические формы). При неблагоприятных условиях у отдельных представителей зооспоры могут функционировать как гаметы.
Половой процесс изо-, гетеро- или оогамный, а у некоторых видов он осуществляется по типу соматогамии. В последнем случае в половом процессе участвуют две соматические (вегетативные) клетки, у которых не происходит дифференциации их содержимого на гаметы. У примитивных хитридиевых грибов соматогамия протекает в форме хологамии. Гаметы образуются в гаметангиях. Зигота превращается в покоящуюся клетку — цисту, с толстой оболочкой и густой цитоплазмой. Цисты иногда могут образоваться и без полового процесса. После периода покоя циста прорастает зооспорангием.
Большинство хитридиомицетов в вегетативном состоянии гаплоидны. Диплоидная фаза у них представлена зиготой. Для некоторых видов отмечена смена поколений. Существование зооспор у хитридиевых указывает на возможную филогенетическую связь со жгутиковыми.
Большинство хитридиомицетов — паразиты пресноводных и морских водорослей, водных грибов, высших водных растений и беспозвоночных животных, и только незначительная часть развивается сапротрофно на растительных и животных остатках. Многие представители этого класса паразитируют на высших наземных растениях, однако их успешное развитие происходит только при сильном увлажнении почвы.
Класс включает 6 порядков, основными из которых являются: Хитридиальные, Бластокладиальные и Моноблефаридаль- ные, различающиеся главным образам степенью развития таллома и типом полового процесса^
Порядок Хитридиальные (Chytridiales). К данному порядку относятся грибы, вегетативное тело которых представлено голой плазменной массой, округлой или цилиндрической клеткой, покрытой оболочкой, от нее отходит более или менее развитый мицелий — ризомицелий.
Бесполое размножение осуществляется одножгутиковыми зооспорами, формирующимися обычно в тонкостенных зооспорангиях. Известны холо- и эукарпические формы. Примитивность строения хитридиальных обусловлена образом жизни — большинство из них являются внутриклеточными паразитами водных, реже наземных растений. Многие развиваются сапротрофно на растительных и животных остатках.
В порядке выделяют около 80 родов и более 400 видов, широко распространенных в Евразии, Африке и Америке. В Беларуси известно несколько родов и более 10 видов. Наибольший интерес представляют роды ольпидиум и синхитриум.
Род ольпидиум (Olpidium) включает грибы с одноклеточным вегетативным телом в виде амебоида, развивающимся в тканях

растений. Зооспорангии шаровидные, с длинным выводным канальцем, одиночные или по 2 — 12 в клетке растения-хозяина (рис. 2.14). В каждом зооспорангии образуется множество мелких яйцевидных одножгутиковых зооспор, выходящих после созревания наружу через выводное отверстие. Оптимальные условия для развития гриба создаются при избыточном увлажнении почвы.
Известно более 20 видов, распространенных в Евразии и Северной Америке. В Беларуси обнаружено 2 вида. Грибы поражают растения капусты, льна, огурцов, помидоров, клевера, пшеницы и др. Наиболее широко распространенным представителем является О. капустный (О. brassicae) — один из возбудителей заболевания рассады капусты, известного под названием «черная ножка». Покоящейся стадией гриба является бесцветная или светло-желтая циста с толстой бородавчатой оболочкой. Весной она прорастает многочисленными зооспорами. При наличии воды зооспоры прилипают к поверхности стебелька капусты, утрачивают жгутики и переливают свое содержимое в клетку эпидермиса. Отсюда паразит проникает в более глубокие слои коры, где разрастается, становится многоядерным, покрывается оболочкой и вновь превращается в
зооспорангий. Формирующиеся в нем зооспоры поражают новые растения. Такой цикл развития может повторяться неоднократно в течение лета. Продолжительность одного цикла — 5—10 дней. При неблагоприятных условиях зооспоры попарно копулируют, образуя двухжгутиковую зиготу, которая проникает в клетку хозяина, увеличивается, покрывается плотной оболочкой и превращается в цисту.
«Черная ножка» капусты проявляется на рассаде разного возраста в виде потемнения корневой шейки или прикорневой части стебля. Корневая шейка у таких растений размягчается, чернеет, резко утончается и загнивает.
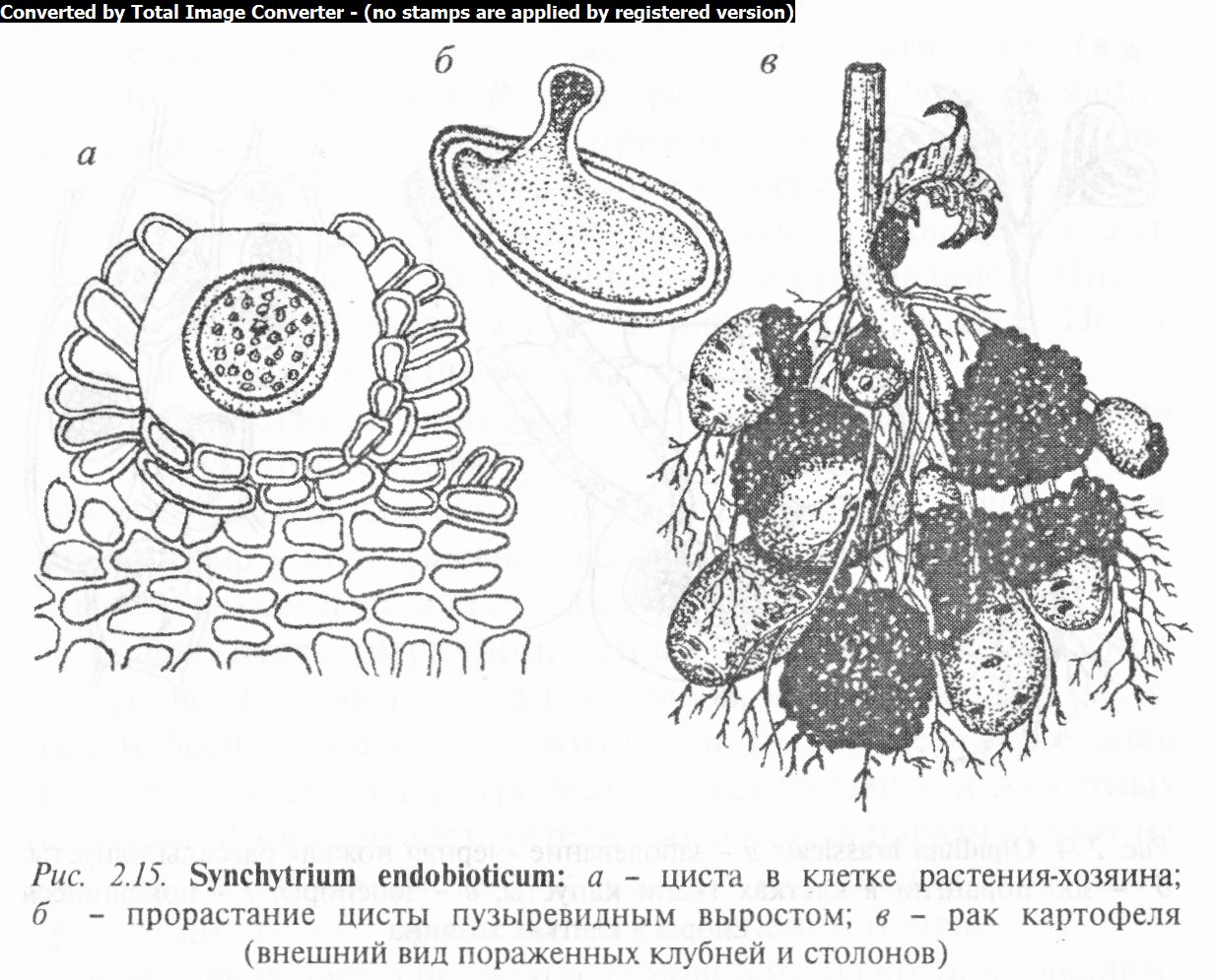
У рода синхитриум (Synchytrium) вегетативное тело (таллом) желтое или красноватое, редко бесцветное, развивается в клетках растений. Позже таллом покрывается тонкой бесцветной оболочкой и превращается в группу зооспорангиев (сорус) или в покоящуюся спору — толстостенную цисту. Покоящиеся споры прорастают пузыревидным выростом, в котором содержимое цисты распадается на отдельные зооспорангии (рис. 2.15, а, б) с бесцветной толстой оболочкой, где формируются многочисленные одножгутиковые зооспоры. Зимуют споры в почве или органах растений. Известно более 200 видов, распространенных на всех материках земного шара. В Беларуси отмечено 5 видов. Часто встречаются С. пролесковый (S. mercuralis), С. ветреницевый (S. anemones), С. одуванчиковый (S. taraxaci). Наиболее интересным представителем является С. внутриклеточный (S. endobioticum), вызывающий болезнь картофеля, известную под названием «рак картофеля».
Неблагоприятные условия этот гриб переносит в виде покоящихся зимних и летних цист. Зимняя циста прорастает в зооспорангии с одножгутиковыми зооспорами, поражающими стебли и листья картофеля и особенно клубни. После завершения вегетации паразит превращается в летние цисты, каждая из которых образует сорус зооспорангиев. Это повторяется в течение лета несколько раз. К концу вегетационного периода зооспоры попарно копулируют с образованием двухжгутиковой планози- готы (подвижной), которая проникает в клетку хозяина и превращается в зимнюю цисту с плотной многослойной оболочкой. Таким образом, в развитии патогена имеются бесполый гаплоидный (зооспоры, амебоид, сорус зооспорангиев) и половой диплоидный (планозигота и зимняя циста) циклы.
На пораженных растениях картофеля появляются вздутия и галлы, а затем происходит деформация органов. Размеры наростов часто превышают величину клубня. Раковые наросты с бугристой темной поверхностью (рис. 2.15, в) легко загнивают и разрушаются, вызывая гниль в здоровой части клубня. Недобор урожая при этой болезни может превышать 50%.
Задания
1. Рассмотреть и зарисовать рассаду капусты, пораженную ольпидиумом. Сделать поперечный срез через место поражения. Рассмотреть под микроскопом и зарисовать возбудителя «черной ножки» на разных стадиях развития — плазмодий, зооспорангии. Цикл развития ольпидиума изобразить схематически.
2. На пораженных грибом Synchytrium endobioticum клубнях картофеля рассмотреть внешнее проявление рака картофеля. Зарисовать общий вид пораженного клубня. Затем кусочек пораженной ткани картофеля поместить на предметное стекло в каплю воды и размять тупым концом препаровальной иглы, накрыть покровным стеклом и рассмотреть при большом увеличении микроскопа. Зарисовать покоящиеся споры - толстостенные цисты. Нарисовать схему цикла развития синхитриума.
Вопросы для самоконтроля
1. Каковы образ жизни и особенности строения вегетативного тела хитридиомицетов?
2. Как размножаются хитридиальные грибы?
3. Охарактеризуйте цикл развития ольпидиума капустного — возбудителя «черной ножки» капусты — и синхитриума внутриклеточного — возбудителя рака картофеля. Каковы меры борьбы с этими болезнями растений?
4. Как можно представить происхождение и эволюцию хитридиальных грибов?
2.8. ОТДЕЛ ЗИГОМИКОТА (ZYGOMYCOTA)
Характерном особенностью грибов данного отдела является полное отсутствие жгутиковой стадии. Вегетативное тело представлено разветвленным многоядерным мицелием, в клеточных стенках которого содержится хитин в комплексе с хитозаном. Мицелий ценотический, иногда достигающий существенной морфологической дифференциации, с тенденцией к образованию многоклеточности. Об этом свидетельствует то, что у части видов в зрелом состоянии образуются клеточные перегородки, разделяющие мицелий на отдельные многоядерные фрагменты. У паразитов беспозвоночных (энтомофторальные и зоопагальные грибы) мицелий изначально многоклеточный.
Бесполое размножение осуществляется путем фрагментации мицелия, неподвижными спорами, развивающимися внутри спорангиев (спорангиоспоры) или на конидиеносцах (конидии). Половой процесс — зигогамия. Основная часть жизненного цикла этих грибов приходится на гаплоидную фазу.
Отдел Зигомикота включает около 600 видов, относящихся к двум классам: Зигомицеты (Zygomycetes) и Трихомицеты (Trichomycetes). Подавляющее большинство их представителей являются наземными организмами. В основном это почвенные сапротрофы, реже паразиты ряда беспозвоночных, грибов, высших растений, теплокровных животных и человека.
Класс Зигомицеты (Zygomycetes)
Грибы этого класса имеют хорошо развитый ценотический мицелий, дифференцированный на столоны и ризоиды. С возрастом их мицелий становится септированным. Жгутиковые
стадии отсутствуют. Бесполое размножение осуществляется спорами или конидиями. Половой процесс — зигогамия, т.е. слияние содержимого двух не дифференцированных на гаметы одно- или многоядерных клеток (гаметангиев). Продукт оплодотворения — зигота, или зигоспора, — покрывается многослойной оболочкой и переходит в состояние покоя. После периода покоя зигоспора прорастает. При этом ее копуляционные диплоидные ядра редукционно делятся и из зигоспоры вырастает росток мицелия, на вершине которого развивается зародышевый спорангий. Внутри такого спорангия формируется множество одноядерных гаплоидных спор, в отличие от многоядер ных спор, возникших бесполым путем. Большинство зигомице- тов — почвенные сапротрофы. Имеются и паразитные формы.
По уровню организации и дифференцировки таллома, особенностям развития, морфологии анаморф и другим признакам класс подразделяют на 6 порядков. Наиболее изученными и важными из них являются мукоральные и энтомофторальные.
Порядок Мукоральные (Mucorales). Мукоральные грибы имеют хорошо развитый несептированный мицелий, достигающий в длину несколько сантиметров. Воздушный мицелий у многих представителей образует столоны — маловетвящиеся гифы с дугообразным ростом: растущая вверх гифа изгибается вниз и на месте ее соприкосновения с субстратом образуется пучок коротких ризоидов, а затем рост гифы продолжается и вырастает новый дугообразный столон. В старых гифах появляются перегородки, по которым такой мицелий может распадаться на отдельные клетки — оидии. Они приобретают шаровидную форму и размножаются почкованием (так называемые мукоровые дрожжи). Изредка образуются хламидоспоры.
Бесполое размножение осуществляется также спорангиоспо- рами и конидиями. Спорангиоспоры образуются в спорангиях, т.е. эндогенно. Типичные спорангии содержат 60 — 100 и более спор и располагаются на приподнятых над субстратом простых или разветвленных спорангиеносцах, которые заканчиваются шаро~, эллипсовидной, конической или грушевидной колонкой. Форма колонки служит важным систематическим признаком.
Другой тип спороношения у этих грибов представлен постепенным переходом от малоспорового спорангия к типичной конидии. Здесь спорангии мелкие, шаровидные, без колонки, с исчезающей оболочкой, содержащие не более 4 — 5 спор. Это так называемые спорангиолы, в которых количество спор может быть доведено до одной. Эта спора срастается с оболочкой спорангиолы и таким образом образуется кони-
дия. У одного и того же вида могут одновременно встречаться спорангии и спорангиолы. В цикле развития мукоральных доминирует бесполое размножение.
Половой процесс — зигогамия с образованием зигоспор.
Большинство мукоральных грибов ведут сапротрофный образ жизни в виде плесени на пищевых продуктах (преимущественно на плодах и овощах, реже на молочных продуктах, мясе, зерне). Некоторые представители являются возбудителями болезней у человека и животных. Ряд видов мукоральных, обладающих высокой ферментативной активностью, используются в пищевой промышленности и приготовлении спиртных напитков.
Порядок включает около 400 видов, распределяющихся в 8 семействах по признаку бесполого размножения. Типичными представителями порядка являются роды пилоболус, мукор, ризопус и др.
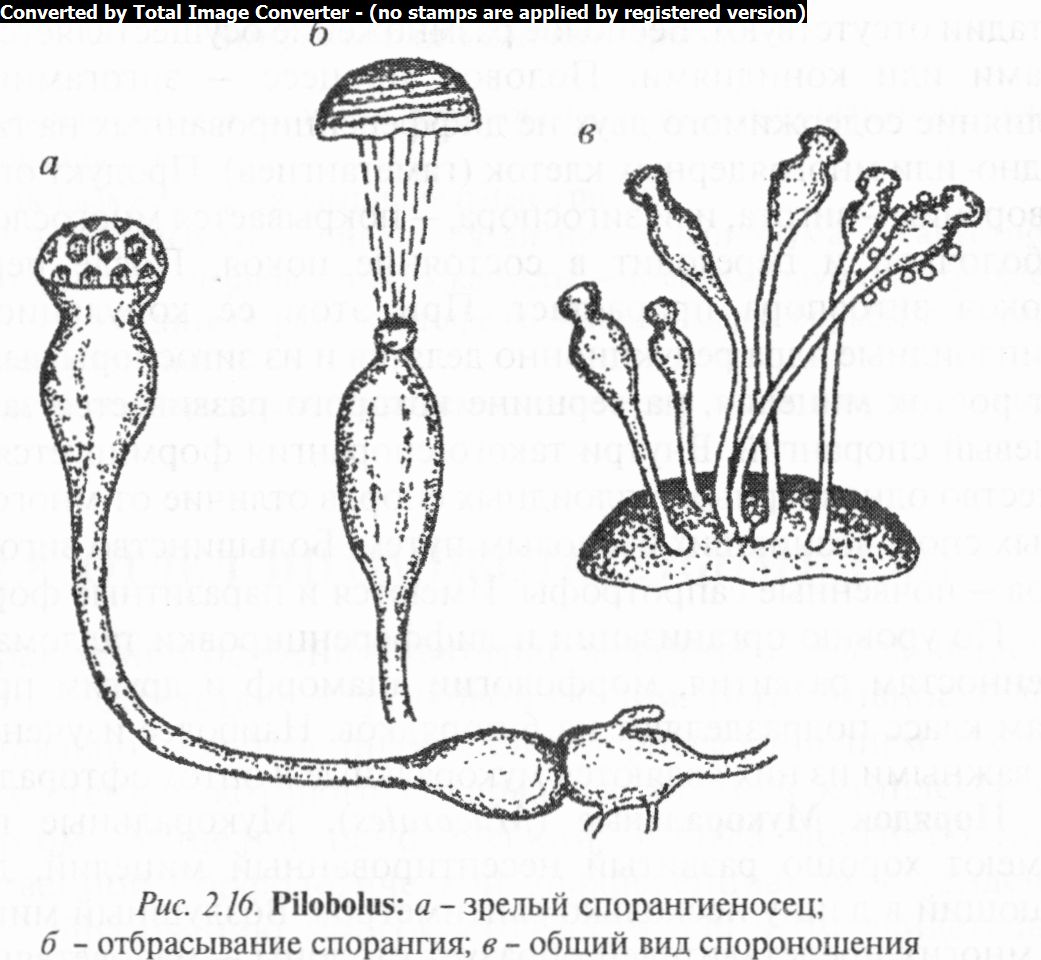
Род пилоболус (Pilobolus) одноименного семейства включает грибы с бесцветным разветвленным мицелием. Короткие (до 10 мм) неветвящиеся спорангиеносцы, вздутые на вершине и оканчивающиеся приплюснутым черным спорангием (рис. 2.16), формируются из особых клеток гиф субстратного мице-
лия — трофоцист. При созревании под действием тургорного давления спорангии с силой отбрасываются на расстояние более 1 м. Приклеившись к траве, спорангии вместе с ней попадают в пищеварительный тракт животных, где разрушаются. Освободившиеся при этом споры вместе с экскрементами выходят наружу и прорастают.
Известно 8 видов в зонах умеренного климата. В Беларуси распространены П. кристаллический (P. crystallimis) и П. рося- нистый (P. roridus), встречающиеся на конском навозе.
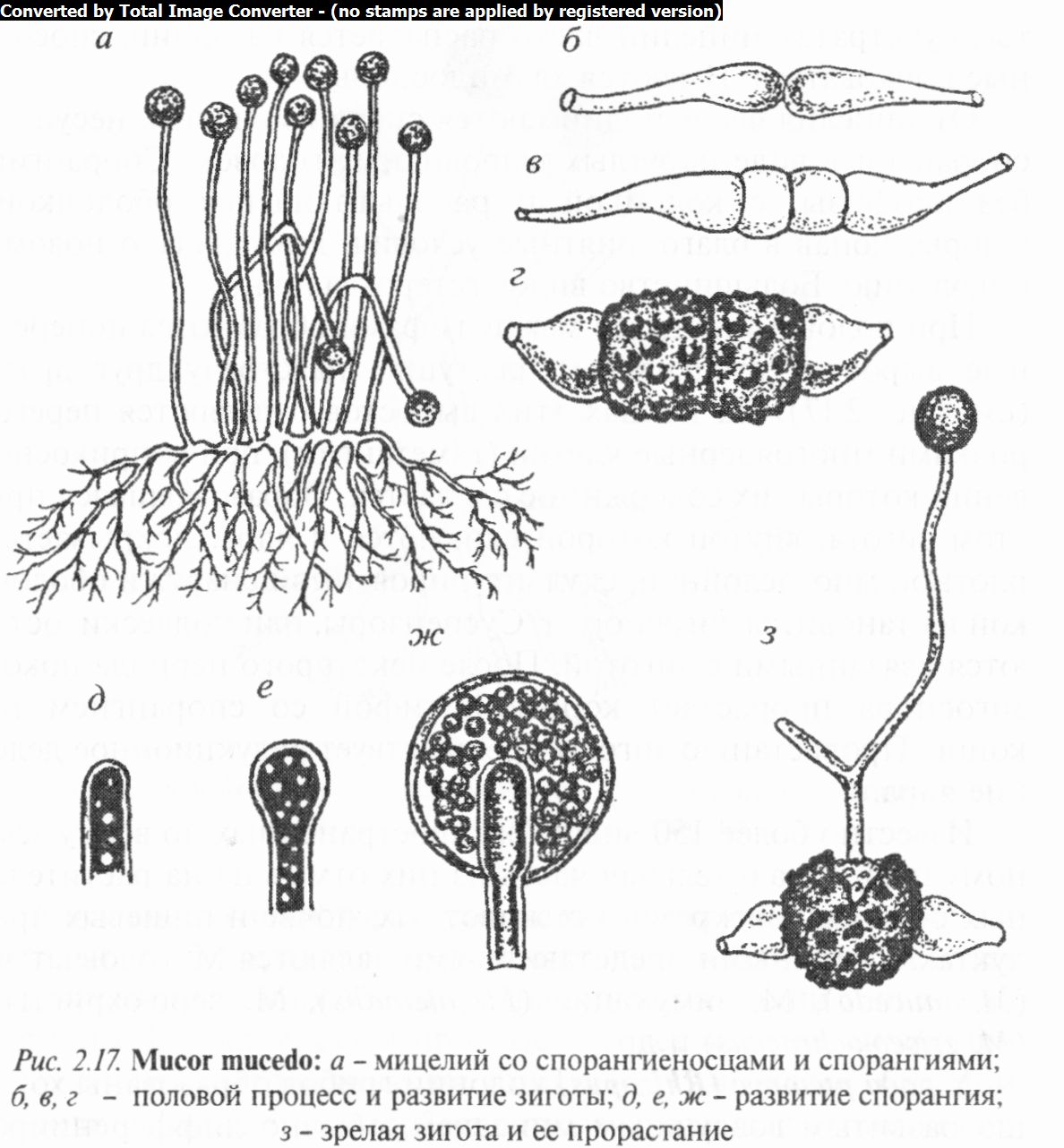
Род мукор (Мисог) одноименного семейства объединяет грибы с хорошо развитым ветвящимся мицелием, пронизывающим субстрат (рис. 2.17). Старые отмершие части мицелия отделяются от последнего перегородками. В жидких сахарис-
тых субстратах мицелий часто распадается на оидии, способные почковаться. Имеются хламидоспоры.
От мицелия вверх поднимаются спорангиеносцы, несущие спорангии в виде округлых шаровидных головок. Спорангии без апофизы, с колонкой и расплывающейся оболочкой. Споры, попав в благоприятные условия, дают начало новому поколению. Большинство видов гетероталличны.
При половом процессе между гифами образуются поперечные выросты (суспензоры), растущие навстречу друг другу (см. рис. 2.17). На концах этих выростов отделяются перегородками многоядерные клетки (гаметангии), при соприкосновении которых их содержимое сливается. Образовавшаяся при этом зигота, внутри которой сливаются и ядра, покрывается плотной многослойной, скульптурированной снаружи оболочкой и становится зигоспорой. Суспензоры, или подвески, остаются связанными с зиготой. После некоторого периода покоя зигоспора прорастает короткой гифой со спорангием на конце. Прорастанию зиготы предшествует редукционное деление ядра.
Известно более 150 видов, распространенных по всему земному шару. Значительная часть из них отмечена на растительных остатках, экскрементах животных, почве и пишевых продуктах. Типичными представителями являются М. головчатый (М. mucedo), М. зимующий (М. hiemalis), М. серо-охристый (М. griseo-ochraceus) и др.
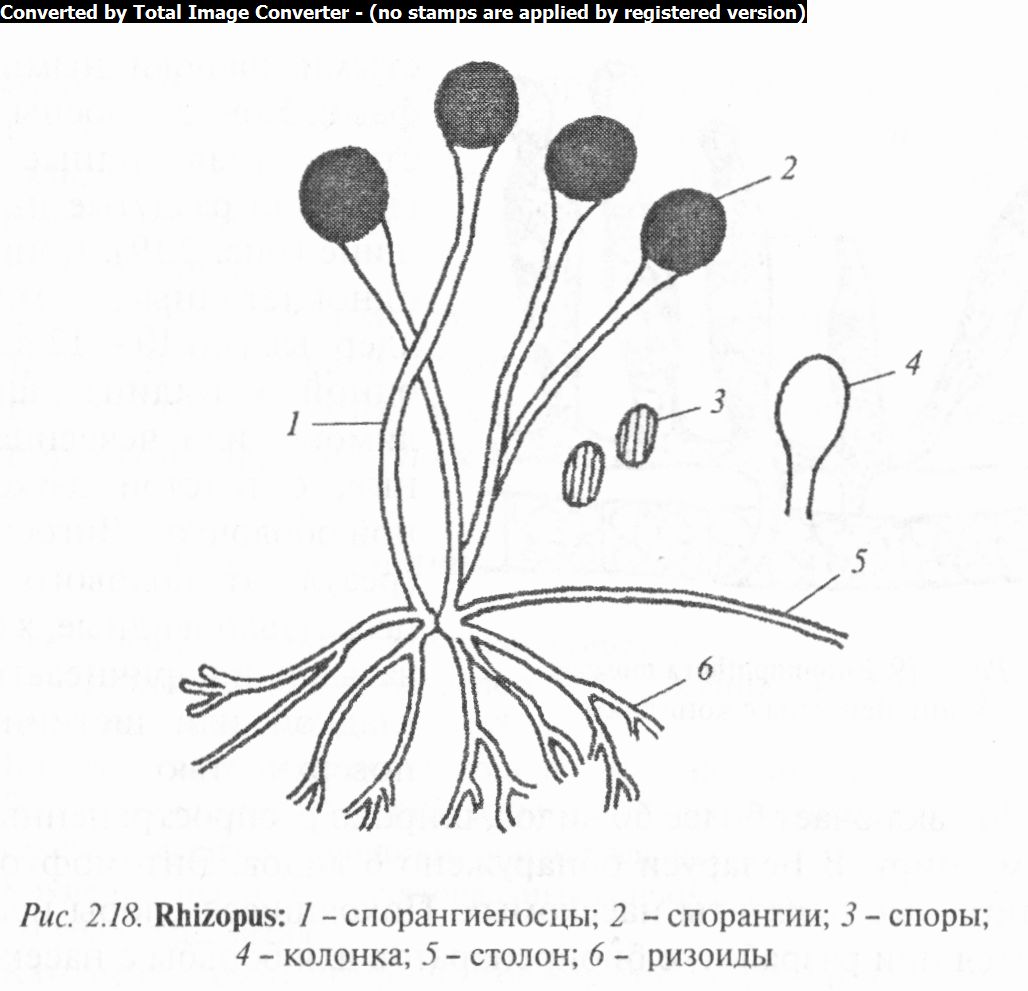
У рода ризопус (Rhizopus) колонии грибов образованы хорошо развитым воздушным мицелием, обычно дифференцированным на столоны, спорангиеносцы и ризоиды. Столоны — толстые, дуговидно изогнутые гифы. Спорангиеносцы простые, расположены на столоне пучками (по 2 — 4). Спорангии крупные, шаровидные, с колонкой, при созревании часто спадающиеся и приобретающие колокольчатую или грибовидную форму (рис. 2.18). Некоторые виды образуют хламидоспоры. Большинство представителей гетероталличны.
Известно 11 видов ризопуса, в том числе в Беларуси 5. Ризо- пусы развиваются сапротрофно, и только некоторые паразитируют. Наиболее распространенный вид Р. черный (Rh. nigricans) — возбудитель черной, или хлебной, плесени. Он часто встречается на заплесневевшем хлебе. Кроме того, он поражает плоды клубники и земляники, что сопровождается отмиранием завязи у цветков, а также может являться причиной мягкой гнили клубней картофеля.
Порядок Энтомофторальные (Entomophthorales). У представителей этого порядка мицелий вначале одноклеточный, затем септированный, способный распадаться на оидии или хлами- доспоры. Конидиеносцы простые или ветвящиеся, цилиндрические или булавовидные, на концах несут по одной конидии. Конидии одноклеточные, бесцветные, гладкие, с двойной оболочкой, при созревании отбрасываются на расстояние до 3 см вследствие разрыва оболочки, оставляющей у вершины кони- диеносца небольшую бахрому в виде воротничка. У некоторых грибов конидии перед прорастанием делятся на несколько частей, каждая из которых дает мицелиальный росток. Половой процесс — зигогамия. Энтомофторальные окончательно потеряли связь с водной средой. Все эти признаки свидетельствуют о высоком уровне развития данной группы грибов и сближают их с аскомицетами.
Порядок Энтомофторальные объединяет одно семейство, 6 родов, около 70 видов. В Беларуси 2 рода и 7 видов. Большинство из них — паразиты насекомых, некоторые сапротрофы на экскрементах лягушек, ящериц, на базидиальных грибах.
Род энтомофтора (Entomophthora) включает виды с обычно внутренним, хорошо развитым мицелием, образованным тол
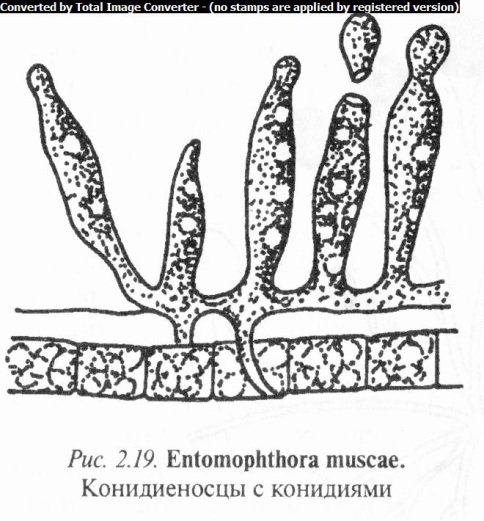
стыми шаровидными гифами. Конидиеносцы простые, булавовидные или внезапно раздутые на вершине (рис. 2.19). Конидии одноклеточные, многоядерные (по 10 — 12 ядер в одной конидии), шаро-, лимоно- или чечевицевид- ные, с толстой двухслойной оболочкой. Зигоспоры (результат полового процесса) шаровидные, желтоватые или коричневатые, с гладкой или щетинистой поверхностью.
Род включает более 60 видов, широко распространенных по всему миру. В Беларуси обнаружено 6 видов. Энтомофтора — облигатные паразиты насекомых. Покоящиеся споры используются при разработке биопрепаратов для борьбы с насекомыми — вредителями сельскохозяйственных культур.
Особенно часто встречаются Э. муховая (Е. muscae), вызывающая «осеннюю болезнь» комнатных мух, Э. скотофагавая (Е'. scatophagae), паразитирующая на навозных мухах, и Э. скученная (Е. conglomerata), паразитирующая на комарах.
Задания
1. Рассмотреть невооруженным глазом и под лупой общий вид плесени, образованной пилоболусом на свежем конском навозе на 5 — 7-й день после помещения его во влажную камеру. Приготовить препарат. Для этого взять пинцетом небольшое количество плесени вместе с кусочком субстрата, перенести в каплю воды на предметном стекле и рассмотреть (без покровного стекла) под бинокуляром или при малом увеличении микроскопа. Рассмотреть и зарисовать часть мицелия, спорангиеносец и спорангий. Обратить внимание на блестящие капельки жидкости, выступающие на поверхности расширенной части спорангиеносца в результате увеличения внутри его тургорного давления.
2.Рассмотреть невооруженным глазом мицелий и спорангиеносцы мукора. Изготовить временный препарат так же, как
и препарат пилоболуса. Изучить при малом и большом увеличении микроскопа и зарисовать важнейшие морфолого-анато- мические особенности мукора. На рисунке обозначить несеп- тированный мицелий, спорангиеносцы, спорангии, споран- гиеспоры, колонку.
3. Изготовить препарат ризопуса таким же способом, как и препараты пилоболуса и мукора. Рассмотреть при малом и большом увеличении микроскопа и зарисовать спорангиеносцы, спорангии, шаровидную колонку, спорангиеспоры.
Вопросы и задания для самоконтроля
1. Дайте общую характеристику зигомицетов.
2. В чем отличие органов бесполого размножения оомицетов и зигомицетов и чем это можно объяснить?
3. Каковы особенности полового процесса у зигомицетов?
4. В чем заключается биологический смысл наблюдающейся тенденции замены спорангиев конидиями у мукоральных?
5. Какие признаки энтомофторальных грибов свидетельствуют об их высоком уровне развития?
6. Каковы распространение и особенности образа жизни мукоральных и энтомофторальных грибов?
7. Охарактеризуйте роль зигомицетов в природе и хозяйственной деятельности человека.
2.9. ОТДЕЛ АСКОМИКОТА (ASCOMYCOTA), ИЛИ СУМЧАТЫЕ ГРИБЫ
Отдел аскомикота включает грибы с хорошо развитым сеп- тированным гаплоидным мицелием. Септы образуются путем инвагинации, т.е. путем впячивания и роста клеточной стенки от периферии к центру. Поэтому в перегородках между клетками имеются поры, через которые перемещаются не только растворенные вещества, но и органеллы. Составными элементами клеточной стенки являются полисахариды хитин и глю- каны.
Характерная особенность грибов этого отдела — образование после полового процесса сумок, или асков, содержащих 4 — 8 (редко 1) аскоспор. Кроме того, диагностическим признаком сумчатых грибов является наличие ламеллярных двуслойных клеточных стенок с тонким электронно-плотным наружным слоем и относительно толстым электронно-прозрач-
ным внутренним слоем. У базидиальных грибов они многослойные.
Вегетативное размножение осуществляется участками мицелия, почкованием или путем расчленения гифы на отдельные клетки и формирования оидий, хламидоспор и т.д.
Важное место в цикле развития аскомицетов играет бесполое размножение при помощи конидий. Отдельные группы грибов могут развивать несколько видов конидиального споро- ношения.
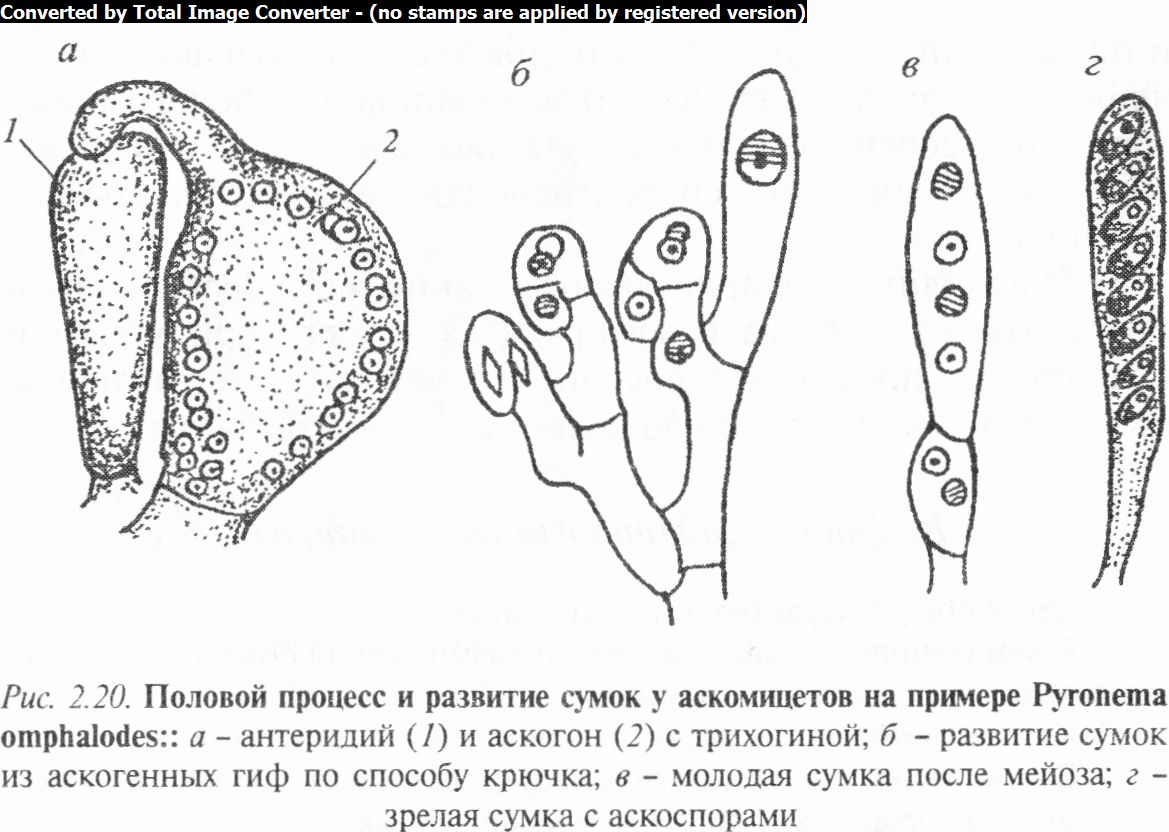
Типы полового процесса определяются уровнем организации гриба. У примитивных, наиболее просто устроенных аско- микот (класс Голосумчатые) половой процесс состоит в слиянии содержимого двух клеток (гаметангиев) мицелия и напоминает собой зигогамию или конъюгацию. Образовавшаяся зигота прямо на мицелии превращается в сумку. При этом копуляционное диплоидное ядро без периода покоя зиготы делится редукционно, затем митотически, образуя 8 гаплоидных ядер, которые облекаются цитоплазмой, формируют оболочку и превращаются таким образом в аскоспоры. Сумки у них прототуникатные, т.е. с тонкой, слабо дифференцированной оболочкой, которая ослизняется или разрушается при созревании аскоспор. Такая сумка не участвует в рассеивании спор, а является лишь их вместилищем.
У высокоорганизованных аскомикот (класс Плодосумча- тые) формируются мужские и женские органы полового размножения - антеридии и архикарпы (рис. 2.20). Архикарп состоит из нижней расширенной части — аскогона и верхней суженной — трихогины. Содержимое антеридия, не дифференцированное на гаметы, перетекает через трихогину в аскогон, где происходит плазмогамия, а гаплоидные ядра разного пола сближаются попарно, образуя дикарионы. Затем аскогон прорастает аскогенными гифами, в которых закладываются поперечные перегородки, образующие клетки с одним или несколькими дикарионами. На концах аскогенных гиф в результате кариогамии с последующим мейозом и митозом образуются сумки с 8 аскоспорами. Сумки возникают внутри или на поверхности плодовых тел, образуемых сплетением гиф мицелия.
Различают 3 типа плодовых тел: 1) клейстотеции — замкнутые плодовые тела обычно с беспорядочно расположенными внутри них сумками: 2) перитеции — имеют отверстие на вершине, а сумки образуются пучком внутри плодовых тел (на дне) и способны по мере созревания активно выбрасывать споры; 3) апотеции — открытые плодовые тела, на поверхности которых палисадным слоем развиваются сумки и бесплодные гифы — парафизы. Совокупность сумок и парафиз составляет гимениальный слой, или гимений. Распространение спор активнее, чем в случае перитеция.
У некоторых грибов формируются ложные плодовые тела — аскостромы, по внешнему виду часто похожие на настоящие перитеции, но отличающиеся от них отсутствием собственного перидия; их оболочкой служит плектенхима стромы.
В цикле развития высокоорганизованных сумчатых грибов закономерно сменяют друг друга 3 ядерные фазы: длительная гаплоидная, представленная долгоживущим мицелием, на котором развивается конидиальное спороношение; кратковременная дикариотическая — от плазмогамии до кариогамии (существует в виде аскогенных гиф) и самая кратковременная диплоидная (материнская клетка сумки) — от кариогамии до мейоза.
Сумчатые грибы — преимущественно сапротрофы на отмерших растительных остатках, почве, пищевых продуктах, различных промышленных материалах и изделиях, вызывают их плесневение и порчу. Некоторые сапротрофы являются продуцентами антибиотиков и других биологически активных веществ. Среди сумчатых т рибов встречаются съедобные виды (сморчки, строчки, трюфели) и паразиты, возбудители болезней растений, животных и человека.
Насчитывают около 30 ООО видов сумчатых грибов, из них в Беларуси отмечено около 500, в том числе 5 видов включены в Красную книгу Республики Беларусь. Сумчатые грибы подразделяются на 4 класса: Археаскомицеты, Гемиаскомицеты, или Голосумчатые, Настоящие сумчатые и Локулоаскомицеты. Систематика отдела основана на расположении сумок: открыто на мицелии, в особых полостях (локулах) или плодовых телах, а также на особенностях их строения.
Класс Археаскомицеты (Archaeascomycetes)
Класс включает одноклеточные и многоклеточные грибы, у которых плодовые тела отсутствуют. Сумки у всех представителей класса эутуникатные, т.е. с плотной оболочкой, часто с особым аппаратом для вскрывания и активного распространения спор.
Класс подразделяют на несколько (от 2 до 5) порядков, основным из которых является порядок Тафринальные.
Порядок Тафринальные (Taplirinales). К этому порядку относятся исключительно паразиты на высших растениях, у которых они вызывают различные деформации пораженных органов (курчавость листьев, «ведьмины метлы», кармашки плодов, галлообразные вздутия и др.).
Настоящий нитчатый мицелий развивается под кутикулой или в межклетниках, однолетний или зимует в почках и побегах, легко распадается на отдельные фрагменты. Сумки образуются пучками или в виде сплошного слоя, выступающего на поверхности пораженного органа. Сумки отходят непосредственно от мицелия или каждая помещается на отдельной подсумочной клетке в виде подставки, отделяющейся от сумки поперечной перегородкой. Сумкоспоры шаро-, эллипсовидные или продолговатые, одноклеточные, бесцветные, по 4 — 8 и более в сумке, часто в ней почкующиеся. Конидиальное споро- ношение у них отсутствует.
В их цикле развития наблюдается длительная дикариотиче- ская фаза, а сумки имеют эутуникатное строение и аскоспоры из них выбрасываются активно. Предполагают, что тафринальные филогенетически связаны с дискомицетами, а отсутствие у них плодовых тел — результат морфологического упрощения в процессе приспособления к паразитизму.
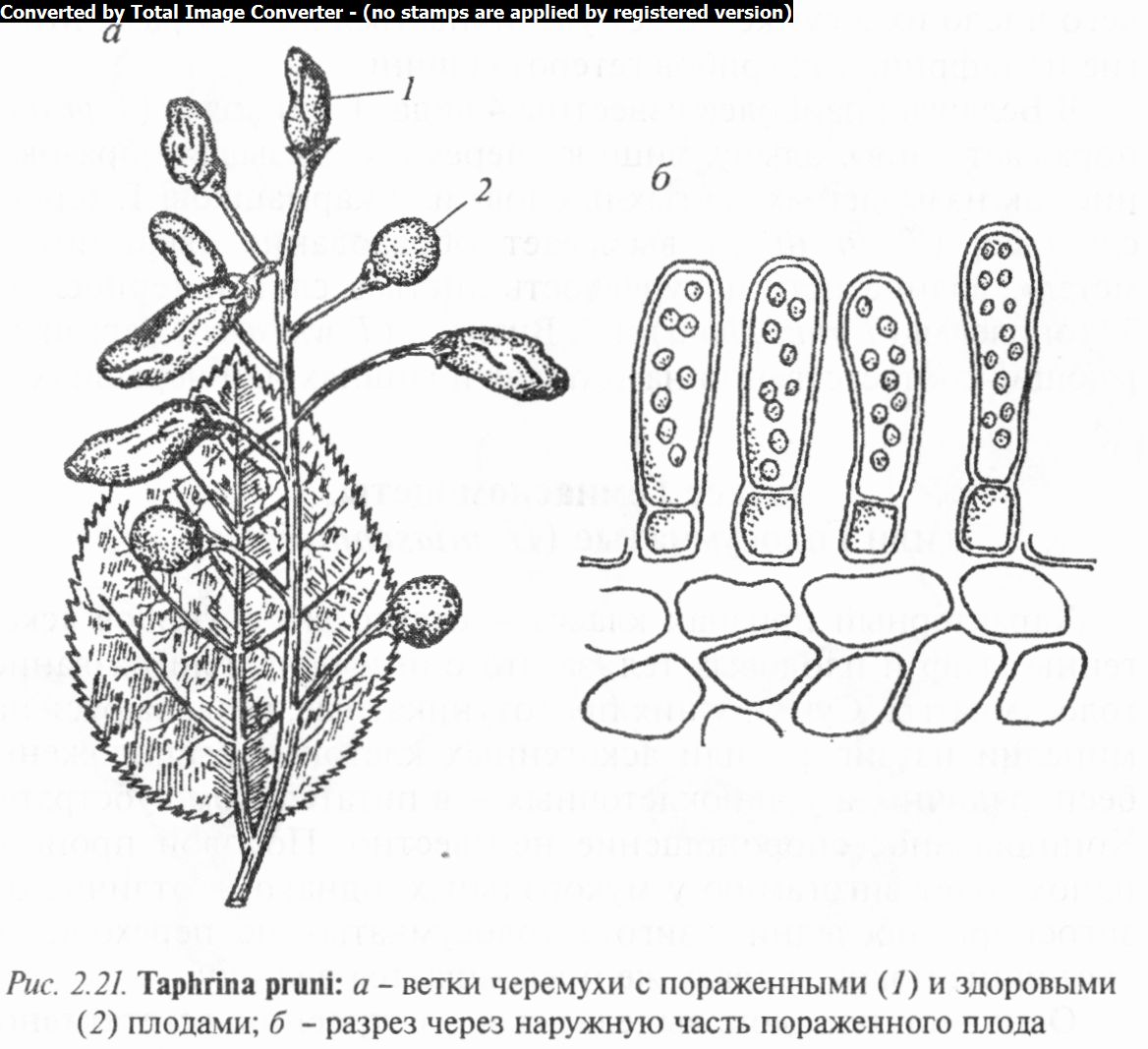
Тафринальные грибы объединены в одно семейство тафри- новых (Taphrinophyceae) с одним родом тафрина, включающим около 100 видов, широко распространенных во многих странах мира Северного и Южного полушарий. В Беларуси встречается около 10 видов, паразитирующих на древесных породах и плодовых культурах, главным образом на косточковых.
Род тафрина (Taphrina) включает грибы с эндофитным мицелием. Сумки образуются непосредственно на мицелии плотным слоем, имеющим сходство с гимением высокоорганизованных сумчатых грибов. Слой сумок обычно имеет желтый, красный, розовый или фиолетовый оттенок. По форме сумки могут быть цилиндрическими, булавовидными или даже удлиненными. Они достигают 60— 110 мкм в высоту и 25 мкм в ширину. В основании сумок находится лишенная протопласта клетка, образовавшаяся в результате деления аскогенной клетки и сохраняющая связь сумки с гифами (рис. 2.21).
В сумке 8 аскоспор округлой или яйцевидной формы, 7 — 10 мкм в диаметре. Они способны к почкованию, в результате
чего число их в сумке может увеличиваться в 2 — 4 раза. Многие из тафриновых грибов гетероталличны.
В Беларуси наиболее известны 4 вида: Т. сливовая (Т. pruni) поражает сливу, алычу, вишню, черемуху, вызывая образование так называемых «дутых плодов» или кармашков; Т. терно- сливовая (Т. insititiae) вызывает образование «ведьминых метел», волнистость и курчавость листьев слив и тернослив; Т. тополевая (Т. rhizophora) и Т. Визнера (Т. wiesneri), паразитирующие соответственно на тополях и вишнях или черешнях.
Класс Гемиаскомицеты, или Голосумчатые (Hemiascomycetes)
Характерный признак класса — отсутствие у грибов аскогенных гиф и плодовых тел, за что они и получили название голосумчатых. Сумки у них прототуникатные, развиваются на мицелии из зиготы или аскогенных клеток и расположены беспорядочно, а у одноклеточных — в питательном субстрате. Конидиальное спороношение неизвестно. Половой процесс напоминает зигогамию у мукоральны<
Дата добавления: 2021-03-18; просмотров: 1110;
Поиск по сайту
Узнать еще
- Class имя_класса : имя_интерфейса
- DirectoryInfo - класс
- DSM — система классификации Американской психиатрической ассоциации
- I. Задачи Единой всероссийской спортивной классификации
- I. История возникновения и развития классно-урочной системы.
- II Классификация САSЕ-средств
- II. Классификация документов
- II.4. Классификация нефтей и газов по их химическим и физическим свойствам

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
