Вакуоли и клеточный сок
Вакуоли содержатся почти во всех растительных клетках. Они представляют собой полости в клетке, заполненные водянистым содержимым – клеточнымсоком. От цитоплазмы клеточный сок изолирован избирательно проницаемой вакуолярной мембраной –тонопластом. Тонопласт выполняет барьерные и транспортные функции.
Для большинства зрелых клеток растений характерна крупная центральная вакуоль, занимающая до 70-90% объема клетки. При этом протопласт со всеми органеллами располагается в виде очень тонкого постенного слоя, выстилающего клеточную стенку. В постенном протопласте обычно встречаются мелкие цитоплазматические вакуоли. Иногда ядро располагается в центре клетки вядерномкармашкецитоплазмы, который связан с постенным слоем тончайшими цитоплазматическими тяжами, пересекающими центральную вакуоль.
Клеточный сок представляет собой водный раствор различных веществ, являющихся продуктами жизнедеятельности протопласта, в основном, запасными веществами и отбросами. Реакция клеточного сока обычно слабокислая или нейтральная, реже щелочная. Вещества, входящие в состав клеточного сока, чрезвычайно разнообразны. Это углеводы, белки, органические кислоты и их соли, аминокислоты, минеральные ионы, алкалоиды, гликозиды, танниды, пигменты и другие растворимые в воде соединения. Большинство из них относится к группе эргастических веществ – продуктов метаболизма протопласта, которые могут появляться и исчезать в различные периоды жизни клетки. Многие вещества клеточного сока образуются только в растительных клетках.
Углеводы клеточного сока растений представлены моносахаридами (глюкоза, фруктоза), дисахаридами (сахароза) и полисахаридами (слизи, инулин).
Глюкоза (виноградный сахар) и фруктоза (плодовый сахар) накапливаются в больших количествах в сочных плодах. Сахароза (свекловичный сахар) в больших количествах накапливается в корнеплодах сахарной свеклы и стеблях сахарного тростника. Для ряда семейств растений (кактусовые, толстянковые, орхидные) характерно накопление в клеточном соке слизей, удерживающих воду. Инулин – запасной полисахарид, откладывается в виде коллоидного раствора в клеточном соке подземных органов сложноцветных вместо крахмала.
Белки накапливаются в виде коллоидного раствора в вакуолях клеток созревающих семян. При обезвоживании семян на поздних этапах их развития вода удаляется из вакуолей, концентрация белка в клеточном соке повышается, и он переходит в состояние твердого геля. Дегидратированные вакуоли зрелых семян называют алейроновымизернами.
Из органических кислот в клеточном соке наиболее часто встречаются лимонная, яблочная, янтарная и щавелевая. Эти кислоты находятся в большом количестве в клеточном соке незрелых плодов, придавая им кислый вкус. При созревании плодов органические кислоты могут использоваться как субстраты дыхания, поэтому кислый вкус плодов обычно исчезает. Соли органических кислот вместе с минеральными ионами играют большую роль в осмотических процессах.
Танниды (дубильные вещества) – полимерные фенольные соединения вяжущего вкуса. Они обладают антисептическими свойствами и защищают ткани растений от инфекций и загнивания. Особенно богаты дубильными веществами клетки коры стеблей и корней (дуб, ива), незрелых плодов (грецкий орех), листьев (чай) и некоторых патологических наростов – галлов. Танниды используются в медицине, для дубления кожи, окраски ткани в темно-коричневый цвет.
Алкалоиды – разнообразные в химическом отношении азотсодержащие органические вещества, имеющие горький вкус. Они обладают свойствами оснований и содержатся в клеточном соке, как правило, в виде солей. Многие алкалоидоносные растения ядовиты и не поедаются травоядными животными. В клетках, содержащих алкалоиды, не развиваются споры и зачатки микроорганизмов, растения не поражаются грибными и бактериальными болезнями. Особенно богаты алкалоидами представители семейств пасленовых, маковых, мареновых, лютиковых и др.
Гликозиды – обширная группа природных веществ, соединения сахаров со спиртами, альдегидами, фенолами и другими веществами. Ряд гликозидов растений используется в медицине. К гликозидам принадлежат также пигменты клеточного сока – флавоноиды. Одни из них –антоцианы– придают клеточному соку красный, синий или фиолетовый цвет; другие –флавоны– желтый. С антоцианами связана окраска цветков многих растений. Цветовая гамма обусловлена реакцией клеточного сока: если она кислая, то господствуют красные тона, нейтральная – фиолетовые, при слабощелочной реакции – синие. На возникновение оттенков оказывает влияние также образование антоцианами комплексов с различными металлами. Флавоны обусловливают желтый цвет лепестков ряда растений.
Значение органических кислот, таннидов, алкалоидов и гликозидов клеточного сока в обмене веществ клетки выяснено недостаточно. Раньше их рассматривали как конечные продукты обмена. В настоящее время показано, что многие из них могут вновь вовлекаться в процессы метаболизма и поэтому их можно рассматривать и как запасные вещества.
Кроме функции накопления запасных веществ и отбросов, вакуоли в растительных клетках выполняют еще одну важную функцию – поддержание тургора. Концентрация ионов и сахаров в клеточном соке центральной вакуоли, как правило, выше, чем в клеточной стенке; тонопласт значительно замедляет диффузию из вакуоли этих веществ и в то же время легко проницаем для воды. Поэтому вода будет поступать в вакуоль. Такой однонаправленный процесс диффузии воды через избирательно проницаемую мембрану носит название осмоса. Поступающая в клеточный сок вода оказывает давление на постенный протопласт, а через него и на клеточную стенку, вызывая напряженное, упругое ее состояние, илитургорклетки. Тургор обеспечивает сохранение неодревесневшими органами растения формы и положения в пространстве, а также их сопротивление действию механических факторов.
Если клетку поместить в гипертонический раствор какой-нибудь нетоксичной соли или сахара (т. е. в раствор большей концентрации, чем концентрация клеточного сока), то происходит осмотический выход воды из вакуоли. В результате этого ее объем сокращается, эластичный постенный протопласт отходит от клеточной стенки, тургор исчезает, наступает плазмолизклетки(рис.2.9).
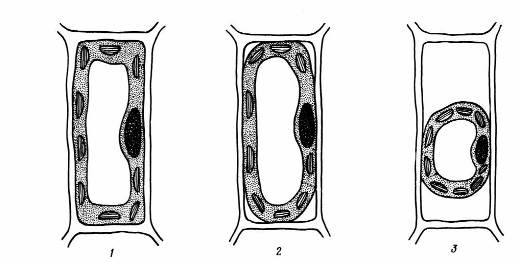
Рис. 2.9.Схема плазмолиза: 1 – клетка в состоянии тургора; 2 – начало плазмолиза; 3 – полный плазмолиз.
Плазмолиз обычно обратим. При помещении клетки в воду или в гипотонический раствор вода снова энергично поглощается центральной вакуолью, протопласт опять прижимается к клеточной стенке, тургор восстанавливается. Плазмолиз может служить показателем живого состояния клетки, мертвая клетка не плазмолизируется, так как не имеет избирательно проницаемых мембран.
Потеря тургора вызывает завядание растения. При завядании на воздухе в условиях недостаточного водоснабжения тонкие стенки клеток сморщиваются одновременно с протопластом и делаются складчатыми.
Тургорное давление не только поддерживает форму неодревесневших частей растений, оно является также одним из факторов роста клетки, обеспечивая ростклетокрастяжением, т. е. за счет поглощения воды и увеличения размера вакуоли. У животных клеток центральная вакуоль отсутствует, их рост происходит главным образом за счет увеличения количества цитоплазмы, поэтому размер животных клеток обычно меньше, чем растительных.
Центральная вакуоль возникает путем слияния многочисленных мелких вакуолей, которые имеются в меристематических (эмбриональных) клетках. Эти цитоплазматические вакуоли образуются, как считают, за счет мембран эндоплазматической сети или аппарата Гольджи.
Включения
Образование включений вызвано избыточным накоплением некоторых продуктов обмена веществ в определенных участках клетки – в вакуоли, гиалоплазме, различных органеллах, реже в клеточной стенке. Эти вещества часто выпадают в осадок в аморфном виде или в форме кристаллов – включений. Включения имеют определенную форму и хорошо видны в световой микроскоп. По наличию тех или иных включений, их форме и распределению можно отличить одни виды, роды и семейства растений от других, поэтому они часто служат важным диагностическим признаком при анализе лекарственного растительного сырья.
Включения представляют собой либо запасныевещества(временно выведенные из обмена веществ соединения), либоконечныепродуктыобмена. К первой категории включений относятсякрахмальныезерна,липидныекаплииотложениябелков; ко второй –кристаллынекоторых веществ.
Крахмальныезерна– наиболее распространенные включения растительных клеток. Полисахаридкрахмал– основной тип запасных питательных веществ растений. Он является и самым важным соединением, используемым в пищу растительноядными животными. Крахмал зерновок хлебных злаков, клубней картофеля, плодов банана – важнейший источник питания людей. Пшеничная мука состоит из зерен крахмала почти на 75%, в клубнях картофеля крахмал составляет 20-30%. В химическом отношении крахмал представляет собой альфа-1,4-D-глюкан, молекулы имеют вид разветвленных цепей, в крахмальном зерне они располагаются по радиусам.
Крахмальные зерна образуются в строме пластид. В хлоропластах на свету откладываются зерна ассимиляционного(первичного)крахмала, образующиеся при избытке сахаров – продуктов фотосинтеза. Образование осмотически неактивного крахмала предотвращает повышение осмотического давления в хлоропласте. Ночью, когда фотосинтез не происходит, ассимиляционный крахмал с помощью ферментов гидролизуется до сахаров и транспортируется в другие части растения.Запасной(вторичный)крахмалоткладывается в амилопластах клеток различных органов растений (корнях, подземных побегах, семенах) из сахаров, притекающих из фотосинтезирующих клеток. При необходимости запасной крахмал также превращается в сахара.
Образование крахмальных зерен начинается в определенных точках стромы пластиды, называемых образовательнымицентрами. Рост зерна происходит путем последовательного отложения слоев крахмала вокруг образовательного центра. Смежные слои в одном зерне могут иметь различный показатель преломления света, и тогда они видны под микроскопом –слоистыекрахмальные зерна. Расположение слоев может бытьконцентрическим(пшеница) илиэксцентрическим(картофель) (рис. 2.10). Если в амилопласте имеется один образовательный центр, вокруг которого откладываются слои крахмала, то возникаетпростоезерно, если два и более – то образуетсясложноезерно, состоящее как бы из нескольких простых.Полусложноезернообразуется в том случае, если крахмал сначала откладывается вокруг нескольких точек, а затем, после соприкосновения простых зерен, вокруг них возникают общие слои (рис. 2.10).
Форма, размер, количество в амилопласте и строение (положение образовательного центра, слоистость, наличие или отсутствие трещин) крахмальных зерен часто специфичны для вида растения (рис. 2.10). Обычно крахмальные зерна имеют сферическую, яйцевидную или линзовидную форму, однако у картофеля она неправильная. Наиболее крупные зерна (до 100 мкм) характерны для клеток клубней картофеля, в зерновке пшеницы они двух размеров – мелкие (2-9 мкм) и более крупные (30-45 мкм). Для клеток зерновки кукурузы характерны мелкие зерна (5-30 мкм). Сложные крахмальные зерна у риса, овса, гречихи.
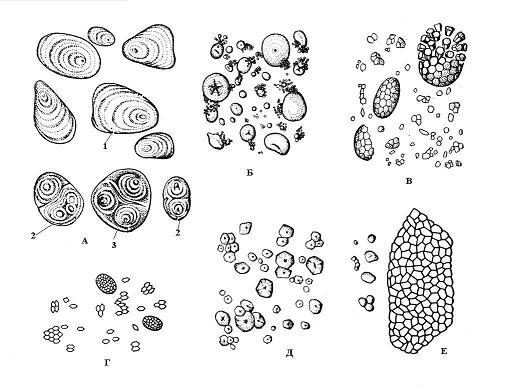
Рис. 2.10.Крахмальные зерна различных видов растений: А – картофель; Б – пшеница; В – овес; Г – рис; Д – кукуруза; Е – гречиха; 1 – простое зерно; 2 – сложное зерно; 3 – полусложное зерно.
Реактивом на крахмал является раствор йода в растворе калия йодида – реактивЛюголя. Он окрашивает крахмальные зерна в сине-фиолетовый цвет.
Отложения крахмала широко распространены во всех органах растения, но особенно богаты им семена, подземные побеги (клубни, луковицы, корневища), паренхима проводящих тканей корней и стеблей древесных растений.
Липидныекапливстречаются практически во всех растительных клетках. Жирные масла накапливаются у огромного количества растений и по своему значению являются второй после крахмала формой запасных питательных веществ. Особенно богаты ими семена и плоды. Семена некоторых растений (подсолнечник, хлопчатник, арахис) могут содержать до 40% масла от массы сухого вещества. Поэтому растительные жиры получают, главным образом, из семян.
Липидные капли накапливаются непосредственно в гиалоплазме. Они выглядят как мелкие сферические тела, каждая капля отделена от гиалоплазмы мембраной. Иногда липидные капли называютсферосомами.
Реактивом на жирное масло является краситель суданIII, липидные капли окрашиваются им в оранжево-красный цвет.
Белковые включенияв виде разнообразных аморфных или кристаллических отложений образуются в различных органеллах клетки. Наиболее часто белковые кристаллы можно встретить в ядре, реже - в гиалоплазме, строме пластид, в расширениях цистерн эндоплазматической сети, матриксе пероксисом и митохондрий. Размер белковых кристаллов чаще всего находится за пределами разрешающей способности светового микроскопа.
Запасные белки относятся к категории простых белков – протеинов, в отличие от сложных белков –протеидов, составляющих основу протопласта. В наибольшем количестве они откладываются в запасающей ткани сухих семян в видеалейроновыхзерен, илибелковыхтелец.
Алейроновые зерна обычно имеют сферическую форму и различный размер (0,2-20 мкм). Они окружены мембраной и содержат аморфный белковый матрикс, в который погружены кристаллические включения – один (реже, 2-3) белковый кристалл ромбоэдрической формы и округлые глобоиды(от одного до многих) (рис. 2.11). Глобоиды состоят из фитина (соли инозитгексафосфорной кислоты) и являются местом хранения запасного фосфора. Алейроновые зерна, содержащие кристаллы, называютсложными. Они характерны для запасающих клеток семян масличных растений (лен, подсолнечник, тыква, горчица, клещевина и др.). Реже встречаютсяпростыеалейроновые зерна, не содержащие кристаллов, а только аморфный белок (бобовые, рис, кукуруза) (рис. 2.12).
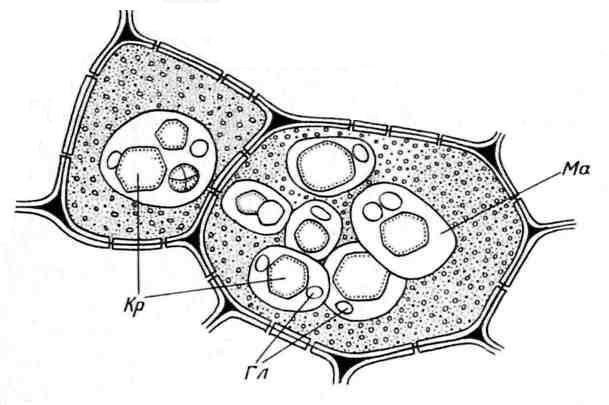
Рис. 2.11.Алейроновые зерна в клетках эндосперма семян клещевины:Кр– белковые кристаллы;Гл– глобоиды;Ма– белковый матрикс.
Запасные белки во время развития семян откладываются в вакуоли. При созревании семян, сопровождающемся их обезвоживанием, белковые вакуоли высыхают, белок и фитин выпадают из раствора в осадок и могут кристаллизоваться. При прорастании семян алейроновые зерна поглощают воду, набухают и постепенно превращаются в типичные вакуоли. Белки и вещества глобоидов расходуются на рост и развитие проростка.
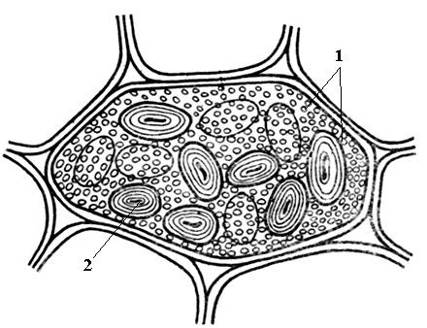
Рис. 2.12.Простые алейроновые и крахмальные зерна в клетке семядоли семени фасоли: 1 – простые алейроновые зерна; 2 – крахмальное зерно.
Белковые включения можно окрасить реактивом Люголя в золотисто-желтый цвет.
Кристаллыкальцияоксалатачасто встречаются в растительных клетках. Они откладываются только в вакуолях. Форма кристаллов кальция оксалата довольно разнообразна (рис. 2.13)и часто специфична для определенных растений, что используется при диагностике лекарственного растительного сырья. Это могут бытьодиночныекристаллыромбоэдрической, октаэдрической или удлиненной формы (листья белены),друзы– звездчатые сростки кристаллов шаровидной формы (листья спорыша, дурмана, сенны, корни ревеня),рафиды– мелкие игольчатые кристаллы, собранные в пучки (листья ландыша, корневища марены),стилоиды– более крупные, палочковидные кристаллы (листья ландыша) икристаллическийпесок– скопления множества мелких одиночных кристаллов (листья красавки). Наиболее часто встречаются друзы. 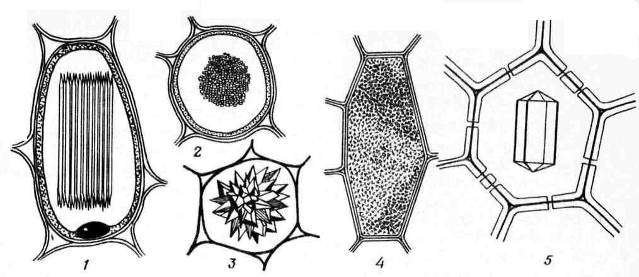
Рис. 2.13.Формы кристаллов кальция оксалата: 1,2 – рафиды (1 – вид сбоку, 2 – вид на поперечном срезе); 3 – друза; 4 – кристаллический песок; 5 – одиночный кристалл.
Вдоль волокон в коре или вдоль жилок листьев у ряда растений (кора дуба, корни солодки, листья сенны) встречается кристаллоносная обкладка– расположенные параллельными рядами клетки с одиночными кристаллами кальция оксалата (рис.2.14).
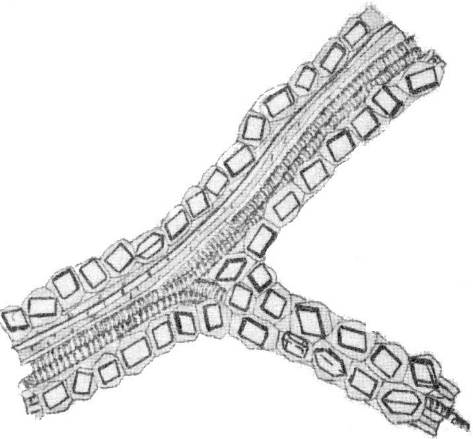
Рис. 2.14.Жилка с кристаллоносной обкладкой в листе сенны.
В отличие от животных, которые выделяют избыток ионов во внешнюю среду вместе с мочой, растения, не имеющие развитых органов выделения, вынуждены накапливать их в тканях. Обычно считают, что кристаллы кальция оксалата – конечный продукт жизнедеятельности клетки, предназначенный для выведения излишков кальция. Действительно, кристаллы образуются в больших количествах в тех органах и тканях, которые растения время от времени сбрасывают (листья и кора). Однако имеются данные, что кристаллы могут исчезать из вакуолей. В таком случае их можно рассматривать как место отложения запасного кальция.
Подтвердить химическую природу кристаллов кальция оксалата можно действием концентрированных минеральных кислот. Под действием кислоты хлористоводородной кристаллы растворяются. При действии кислоты серной кальция оксалат переходит в нерастворимый кальция сульфат (гипс), образующий многочисленные игольчатые кристаллы.
К кристаллическим включениям близки цистолиты. Они чаще всего состоят из кальция карбоната или кремнезема и представляют собой гроздевидные образования, возникающие на выступах клеточной стенки, вдающейся внутрь клетки (рис. 2.15). Цистолиты характерны для растений семейств крапивных, тутовых. Значение цистолитов пока не выяснено.
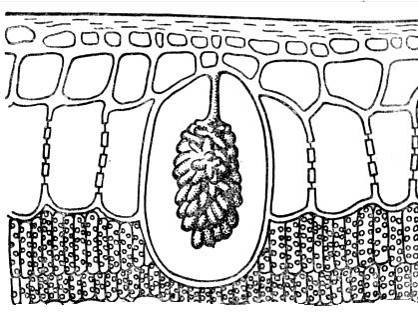
Рис. 2.15.Цистолит в клетке эпидермы листа фикуса.
Клеточная стенка
Клеточная стенка (клеточная оболочка)– характерный признак растительной клетки, отличающий ее от клетки животной. Клеточная стенка придает клетке определенную форму. Культивируемые на специальных питательных средах клетки растений, у которых ферментативным путем удаляется стенка, всегда принимают сферическую форму. Клеточная стенка придает клетке прочность и защищает протопласт, она уравновешивает тургорное давление и препятствует, таким образом, разрыву плазмалеммы. Совокупность клеточных стенок образует внутренний скелет, поддерживающий тело растения и придающий ему механическую прочность.
Клеточная стенка бесцветна и прозрачна, легко пропускает солнечный свет. Обычно стенки пропитаны водой. По системе клеточных стенок осуществляется транспорт воды и растворенных в ней низкомолекулярных соединений (транспорт по апопласту).
Клеточная стенка состоит в основном из полисахаридов, которые можно подразделить на скелетные вещества и вещества матрикса.
Скелетным веществомклеточной стенки растений являетсяцеллюлоза (клетчатка), представляющая собой бета-1,4-D-глюкан. Это самое распространенное органическое вещество биосферы. Молекулы целлюлозы представляют собой очень длинные неразветвленные цепи, они располагаются параллельно друг другу группами по нескольку десятков и скреплены многочисленными водородными связями. В результате образуютсямикрофибриллы, которые создают структурный каркас стенки и обусловливают ее прочность. Микрофибриллы целлюлозы видны только в электронный микроскоп, их диаметр равен 10-30 нм, длина достигает нескольких мкм.
Целлюлоза нерастворима и не набухает в воде. Она очень инертна в химическом отношении, не растворяется в органических растворителях, концентрированных щелочах и разведенных кислотах. Микрофибриллы целлюлозы эластичны и очень прочны на разрыв (сходны со сталью). Эти свойства определяют широкое применение целлюлозы и ее продуктов. Мировая продукция хлопкового волокна, состоящего почти целиком из целлюлозы, составляет 1,5•107 тонн в год. Из целлюлозы получают бездымный порох, ацетатный шелк и вискозу, целлофан, бумагу. Качественную реакцию на целлюлозу проводят с реактивомхлор-цинк-йод, целлюлозная клеточная стенка окрашивается в сине-фиолетовый цвет.
У грибов скелетным веществом клеточной стенки является хитин – полисахарид, построенный из остатков глюкозамина. Хитин еще более прочен, чем целлюлоза.
Микрофибриллы погружены в аморфный матрикс, обычно представляющий собой насыщенный водой пластичный гель. Матрикс является сложной смесью полисахаридов, молекулы которых состоят из остатков нескольких различных сахаров и представляют собой более короткие, чем у целлюлозы, и разветвленные цепи. Матричные полисахариды определяют такие свойства клеточной стенки, как сильная набухаемость, высокая проницаемость для воды и растворенных в ней низкомолекулярных соединений, катионообменные свойства. Полисахариды матрикса делят на две группы –пектиновые вещества игемицеллюлозы.
Пектиновые веществасильно набухают или растворяются в воде. Они легко разрушаются под действием щелочей и кислот. Простейшими представителями пектиновых веществ являются растворимые в водепектовые кислоты – продукты полимеризации альфа-D-галактуроновой кислоты (до 100 единиц), связанных 1,4-связями в линейные цепи (альфа-1,4-D-галактуронан).Пектиновые кислоты (пектины)– это более высокомолекулярные (100-200 единиц) полимерные соединения альфа-D-галактуроновой кислоты, в которых карбоксильные группы частично метилированы.Пектатыипектинаты– кальциевые и магниевые соли пектовых и пектиновых кислот. Пектиновые кислоты, пектаты и пектинаты растворимы в воде в присутствии сахаров и органических кислот с образованием плотных гелей.
В клеточных стенках растений в основном присутствуют протопектины – высокомолекулярные полимеры метоксилированной полигалактуроновой кислоты с арабинанами и галактанами, у двудольных растений в состав цепей галактуронана входит небольшое количество рамнозы. Протопектины нерастворимы в воде.
Гемицеллюлозыпредставляют собой разветвленные цепи, построенные из остатков нейтральных сахаров, чаще встречаются глюкоза, галактоза, манноза, ксилоза; степень полимеризации 50-300. Гемицеллюлозы химически более устойчивы, чем пектиновые вещества, они труднее гидролизуются и слабее набухают в воде. Гемицеллюлозы могут откладываться в стенках клеток семян в качестве запасных веществ (финиковая пальма, хурма). Пектиновые вещества и гемицеллюлозы связаны взаимными переходами. Помимо полисахаридов, в матриксе клеточных стенок присутствует особый структурный белок. Он связан с остатками сахара арабинозы и поэтому является гликопротеидом.
Матричные полисахариды не просто заполняют промежутки между целлюлозными микрофибриллами. Их цепи располагаются упорядоченно и образуют многочисленные связи как друг с другом, так и с микрофибриллами, что значительно повышает прочность клеточной стенки.
Клеточные стенки растений часто подвергаются химическим видоизменениям. Одревеснение, илилигнификацияпроисходит в том случае, если в матриксе откладываетсялигнин – полимерное соединение фенольной природы, нерастворимое в воде. Одревесневшая клеточная стенка теряет эластичность, резко повышается ее твердость и прочность на сжатие, снижается проницаемость для воды. Реактивами на лигнин являются: 1)флороглюцин иконцентрированная хлористоводороднаяилисерная кислота(одревесневшие стенки приобретают вишнево-красную окраску) и 2)сульфатанилина, под действием которого одревесневшие стенки становятся лимонно-желтыми. Лигнификация характерна для стенок клеток проводящей ткани ксилемы (древесины) и механической ткани склеренхимы.
Опробковение, илисуберинизацияпроисходит в результате отложения с внутренней стороны клеточной стенки гидрофобных полимеров –суберинаивоска. Суберин представляет собой смесь эфиров полимерных жирных кислот. Мономерами воска являются жирные спирты и восковые эфиры. Воск легко извлекается органическими растворителями и быстро плавится, образует кристаллы. Суберин – аморфное соединение, не плавится и не растворяется в органических растворителях. Суберин и воск, образуя чередующиеся параллельные слои, выстилают всю полость клетки с внутренней стороны в виде пленки. Субериновая пленка практически непроницаема для воды и для газов, поэтому после ее образования клетка обычно отмирает. Опробковение характерно для стенок клеток покровной ткани пробки. Реактивом на опробковевшую клеточную стенку являетсясудан III, окраска оранжево-красная.
Кутинизацииподвергаются наружные стенки клеток покровной ткани эпидермы.Кутинивоскоткладываются чередующимися слоями на наружной поверхности клеточной стенки в виде пленки –кутикулы. Кутин представляет собой жироподобное полимерное соединение, близкое по химической природе и свойствам суберину. Кутикула предохраняет растение от излишнего испарения воды с поверхности растения. Окрасить ее можно реактивомсудан IIIв оранжево-красный цвет.
Минерализацияклеточной стенки происходит вследствие отложения в матриксе большого количества минеральных веществ, чаще всего кремнезема (оксида кремния), реже оксалата и карбоната кальция. Минеральные вещества придают стенке твердость и хрупкость. Отложение кремнезема характерно для клеток эпидермы хвощей, осок и злаков. Приобретенная в результате окремнения жесткость стеблей и листьев служит защитным средством против улиток, а также значительно снижает поедаемость и кормовую ценность растений.
У некоторых специализированных клеток наблюдается ослизнениеклеточной стенки. При этом вместо целлюлозной вторичной стенки происходит отложение аморфных, сильно гидратированных кислых полисахаридов в видеслизейикамедей, близких по химической природе к пектиновым веществам. Слизи хорошо растворяются в воде с образованием слизистых растворов. Камеди клейкие, вытягиваются в нити. В сухом виде они имеют роговую консистенцию. При отложении слизи протопласт постепенно оттесняется к центру клетки, его объем и объем вакуоли постепенно уменьшаются. В конце концов, полость клетки может целиком заполниться слизью, и клетка отмирает. В некоторых случаях слизь может проходить через первичную клеточную стенку на поверхность. В синтезе и секреции слизи основное участие принимает аппарат Гольджи.
Выделяемая растительными клетками слизь выполняет различные функции. Так, слизь корневого чехлика служит в качестве смазки, облегчающей рост кончика корня в почве. Слизевые железки насекомоядных растений (росянка) выделяют ловчую слизь, к которой приклеиваются насекомые. Слизь, выделяемая наружными клетками семенной кожуры (лен, айва, подорожники), закрепляет семя на поверхности почвы и защищает проросток от высыхания. Слизь окрашивается реактивом метиленовый синийв голубой цвет.
Выделение камедей обычно происходит при поранении растений. Например, камедетечение из пораненных участков стволов и ветвей часто наблюдается у вишни и сливы. Вишневый клей представляет собой застывшую камедь. Камедь выполняет защитную функцию, закрывая рану с поверхности. Образуются камеди в основном у древесных растений из семейств бобовых (акации, трагакантовые астрагалы) и розоцветных подсемейства сливовых (вишня, слива, абрикос). Камеди и слизи используются в медицине.
Клеточная стенка является продуктом жизнедеятельности протопласта. Полисахариды матрикса, гликопротеид стенки, лигнин и слизи образуются в аппарате Гольджи. Синтез целлюлозы, образование и ориентация микрофибрилл осуществляются плазмалеммой. Большая роль в ориентации микрофибрилл принадлежит микротрубочкам, которые располагаются параллельно откладывающимся микрофибриллам вблизи плазмалеммы. Если микротрубочки разрушить, образуются только изодиаметрические клетки.
Образование клеточной стенки начинается во время деления клетки. В плоскости деления образуется клеточная пластинка, единый слой, общий для двух дочерних клеток. Она состоит из пектиновых веществ, имеющих полужидкую консистенцию; целлюлоза отсутствует. Во взрослой клетке клеточная пластинка сохраняется, но претерпевает изменения, поэтому ее называют срединной, илимежклеточной пластинкой(межклеточным веществом)(рис. 2.16). Срединная пластинка обычно очень тонка и почти неразличима.
Сразу после образования клеточной пластинки протопласты дочерних клеток начинают откладывать собственную клеточную стенку. Она откладывается изнутри как на поверхности клеточной пластинки, так и на поверхности других клеточных стенок, принадлежавших ранее материнской клетке. После деления клетка вступает в фазу роста растяжением, который обусловлен интенсивным осмотическим поглощением клеткой воды, связанным с образованием и ростом центральной вакуоли. Тургорное давление начинает растягивать стенку, но она не рвется благодаря тому, что в нее постоянно откладываются новые порции микрофибрилл и веществ матрикса. Отложение новых порций материала происходит равномерно по всей поверхности протопласта, поэтому толщина клеточной стенки не уменьшается.
Стенки делящихся и растущих клеток называют первичными. Они содержат много (60-90%) воды. В сухом веществе преобладают матричные полисахариды (60-70%), содержание целлюлозы не превышает 30%, лигнин отсутствует. Толщина первичной стенки очень невелика (0,1-0,5 мкм).
Для многих клеток отложение клеточной стенки прекращается одновременно с прекращением роста клетки. Такие клетки окружены тонкой первичной стенкой до конца жизни (рис. 2.16).
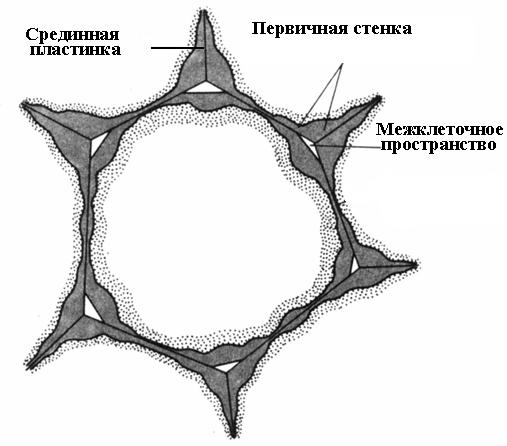
Рис. 2.16.Паренхимная клетка с первичной стенкой.
У других клеток отложение стенки продолжается и по достижении клеткой окончательного размера. При этом толщина стенки увеличивается, а объем, занимаемый полостью клетки, сокращается. Такой процесс носит название вторичного утолщениястенки, а саму стенку называютвторичной(рис. 2.17).
Вторичная стенка может рассматриваться как дополнительная, выполняющая главным образом механическую, опорную функцию. Именно вторичная стенка ответственна за свойства древесины, текстильного волокна, бумаги. Вторичная стенка содержит значительно меньше воды, чем первичная; в ней преобладают микрофибриллы целлюлозы (40-50% от массы сухого вещества), которые располагаются параллельно друг другу. Из полисахаридов матрикса характерны гемицеллюлозы (20-30%), пектиновых веществ очень мало. Вторичные клеточные стенки, как правило, подвергаются одревеснению. В неодревесневших вторичных стенках (лубяные волокна льна, волоски хлопчатника) содержание целлюлозы может достигать 95%. Большое содержание и строго упорядоченная ориентация микрофибрилл определяют высокие механические свойства вторичных стенок. Часто клетки, имеющие вторичную одревесневшую клеточную стенку, после завершения вторичного утолщения отмирают.
Срединная пластинка склеивает соседние клетки. Если ее растворить, стенки клеток теряют связь друг с другом и разъединяются. Этот процесс называется мацерация.Довольно обычна естественная мацерация, при которой пектиновые вещества срединной пластинки переводятся в растворимое состояние с помощью фермента пектиназы и затем вымываются водой (перезрелые плоды груши, дыни, персика, банана). Часто наблюдается частичная мацерация, при которой срединная пластинка растворяется не по всей поверхности, а лишь в углах клеток. Вследствие тургорного давления соседние клетки в этих местах округляются, в результате чего образуютсямежклетники(рис. 2.16). Межклетники образуют единую разветвленную сеть, которая заполняется парами воды и газами. Таким образом, межклетники улучшают газообмен клеток.
Характерная особенность вторичной стенки – ее неравномерное отложение поверх первичной стенки, в результате чего во вторичной стенке остаются неутолщенные участки – поры. Если вторичная стенка не достигает большой толщины, поры выглядят как мелкие углубления. У клеток с мощной вторичной стенкой поры в разрезе имеют вид радиальных каналов, идущих от полости клетки до первичной стенки. По форме порового канала различают поры двух типов –простые и окаймленные (рис. 2.17).
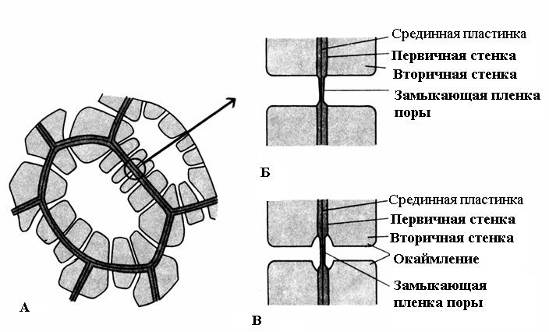
Рис. 2.17.Типы пор: А – клетки с вторичными стенками и многочисленными простыми порами; Б – пара простых пор; В – пара окаймленных пор.
У простых пордиаметр порового канала по всей длине одинаковый и имеет форму узкого цилиндра. Простые поры характерны для паренхимных клеток, лубяных и древесинных волокон.
Поры в двух смежных клетках, как правило, возникают друг против друга. Эти общие поры имеют вид одного канала, разделенного тонкой перегородкой из срединной пластинки и первичной стенки. Такая совокупность двух пор смежных стенок соседних клеток носит название пары пори функционирует как одно целое. Разделяющий их канал участок стенки называетсязамыкающей пленкойпоры, илипоровой мембраной. В живых клетках замыкающая пленка поры пронизана многочисленнымиплазмодесмами (рис. 2.18).
Плазмодесмыприсущи только растительным клеткам. Они представляют собой тяжи цитоплазмы, пересекающие стенку смежных клеток. Число плазмодесм в одной клетке очень велико – от нескольких сотен до десятков тысяч, обычно плазмодесмы собраны в группы. Диаметр плазмодесменного канала составляет 30-60 нм. Его стенки выстланы плазмалеммой, непрерывной с плазмалеммой смежных клеток. В центре плазмодесмы проходит мембранный цилиндр –центральный стерженьплазмодесмы, непрерывный с мембранами элементов эндоплазматической сети обеих клеток. Между центральным стержнем и плазмалеммой в канале находится гиалоплазма, непрерывная с гиалоплазмой смежных клеток.
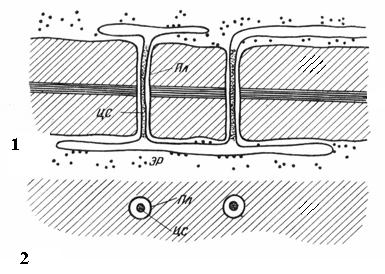
Рис. 2.18.Плазмодесмы под электронным микроскопом (схема): 1 – на продольном срезе; 2 – на поперечном срезе;Пл – плазмалемма;ЦС– центральный стержень плазмодесмы;ЭР– элемент эндоплазматического ретикулума.
Таким образом, протопласты клеток не полностью изолированы друг от друга, а сообщаются по каналам плазмодесм. По ним происходит межклеточный транспорт ионов и мелких молекул, а также передаются гормональные стимулы. Посредством плазмодесм протопласты клеток в растительном организме образуют единое целое, называемое симпластом, а транспорт веществ через плазмодесмы получил названиесимпластическогов отличие отапопластическоготранспорта по клеточным стенкам и межклетникам.
У окаймленных пор(рис. 2.17)канал резко суживается в процессе отложения клеточной стенки, поэтому внутреннее отверстие поры, выходящее в полость клетки, гораздо уже, чем наружное, упирающееся в первичную стенку. Окаймленные поры характерны для рано отмирающих клеток водопроводящих элементов древесины. У них поровый канал по направлению к замыкающей пленке воронковидно расширяется, а вторичная стенка нависает в виде валика над расширенной частью канала, образуя камеру поры. Название окаймленной поры происходит оттого, что при рассмотрении с поверхности внутреннее отверстие имеет вид маленького круга или узкой щели, тогда как наружное отверстие как бы окаймляет внутреннее в виде круга большего диаметра или более широкой щели.
Поры облегчают транспорт в
Дата добавления: 2016-05-30; просмотров: 10849;
Поиск по сайту
Узнать еще
- V.3. Аномально высокие и аномально низкие пластовые давления (АВПД и АНПД).
- А) Совместная работа элементов турбокомпрессора высокого давления.
- Адаптация к действию высокой температуры
- Адаптивный высокочастотный фильтр
- Арматура маслонаполненных кабельных линий высокого давления
- Ассортимент свечей зажигания и высоковольтных проводов в АВТО 49
- Астрономия в эпоху досократиков (и кое-то из ранних цивилизаций)
- Атрад Сокалападобныя

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
