Тип Хордовые – Chordata.
Нервная система хордовых животных
Завершающий и наиболее совершенный тип в царстве животных.
В состав типа входит 3 подтипа: Бесчерепные – Acrania с одним классом Головохордовые – Cephalochordata; Оболочники, или Личиночнохордовые – Tunicata seu Urochordata c 3 классами: Ascidiae, Salpae, Appendiculariae; Черепные или Позвоночные – Craniota seu Vertebrata.
Классы: Круглоротые – Cyclostomata, Хрящевые рыбы – Chondrichthyes, Костные рыбы – Osteichthyes, Земноводные – Amphibia, Пресмыкающиеся – Reptilia, Птицы – Aves, Млекопитающие (Звери) – Mammalia seu Theria. Бесчерепные и Оболочники относятся к низшим хордовым, а Позвоночные к высшим хордовым, составляющие основную массу из примерно 43 тысяч видов хордовых. Позвоночные животные распространены по всему Земному шару и заселяют все арены жизни.
Позвоночные широко распространены во всех сферах Земли. Обитатели глубин мирового океана и заоблачных вершин, полярных льдов и пустынь, они заселяют все возможные экологические ниши, будучи в совершенстве приспособленными к обитанию в тех или иных условиях среды.
Усложнение строения организмов хордовых, совершенство структур, высокий уровень адаптации к среде, естественно, происходили на фоне усиления регуляции и координации процессов жизнедеятельности посредством центральной нервной системы и функционально связанной с ней гормональной регуляцией. По сравнению с беспозвоночными, обе системы отличаются большой сложностью. Именно, совокупность нервных и гормональных механизмов обеспечивают возможность организмов постоянно приспосабливаться к нестабильной окружающей среде при сохранении устойчивости среды внутренней. Механизмы нервной регуляции работают в высоком режиме, так что ответ на раздражение может происходить за доли секунды. Такая реактивность – необходимое условие жизненности высокоподвижных организмов, обитающих в сложных и разнообразных условиях.
Хордовым свойственен совершенно особый тип нервной системы, и в первуюочередь – центральной. Именно трубчатая форма центральной нервной системы является одним из наиболее характерных признаков животных этого класса. Вся центральная нервная система лежит со спинной стороны тела.
У позвоночных передний конец трубки расширен и образует головной мозг. Задняя, цилиндрическая часть трубки являет собой спинной мозг. Число нервных клеток головного мозга огромно и различно в разных классах (более миллиарда у млекопитающих). Вместе с рецепторами центральная нервная система создает сложные нейронные цепи, которые обеспечивают сбор, передачу и переработку информации о состоянии организма и окружающей среды, формируют и передают команды, по которым осуществляются ответные реакции организма.
С развитием центральной нервной системы усложняются орган чувств. И если у низших хордовых число их ограничено и они просты по своей организации, то у высших хордовых позвоночных, имеются многие нервные структуры сложного устройства.
Этапы формирования нервной системы хордовых животных
Нервная система у представителей хордовых, в равной мере как у низкоорганизованного ланцетника, так и у позвоночных на ранних стадиях развития закладывается из однородных структур и морфологически сходна.
В процессе первичного органогенеза, то есть на этапе образования из оплодотворенного яйца гаструлы, а затем с зародышевых листков – экто- и энтодермы, начинается формирование зародыша (рис. 37).
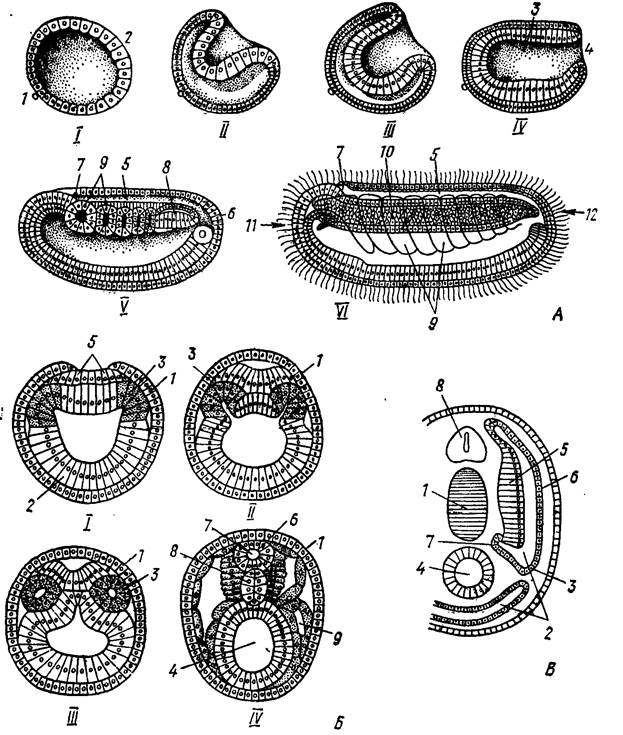

Рис. 37. Эмбриональное развитие хордовых животных (на примере ланцетника) (по Н. П. Наумову, Н. Н. Карташеву, 1979).
В вытянувшейся в горизонтальном направлении гаструле уже пpeдполагается передний "головной" и задний край. В равной мере определяется и спинная сторона, сложенная из мелких эктодермальных клеток анимального полюса бластулы. В зоне гастральной полости из энтодермальных клеток начинают закладываться хорда, кишка. Одновременно на спинной стороне зародыша из клеток упомянутых клеток эктодермы начинают выделяться среди окружающих клеток продолговатая, тянущаяся вдоль тела зародыша нервная пластинка, полоса утолщенной эктодермы. Будучи ограничена с боков невысокими валиками, она погружается под кожу и валики срастаются над ней. В процессе роста как в длину, так и в ширину, нервная пластинка заворачивается с боков образуя нервную трубку с полостью – невроцелем. Передний головной край трубки на эмбриональном уровне имеет наружное отверстие – невропор, задний посредством первичной кишки. По завершении стадии зародыша оба эти отверстия закрываются. На следующем этапе нервная трубка дифференцируется на головной и спинной мозг. В ее передней части возникает небольшое вздутие, из которого последовательно образуются три мозговых пузыря: передний, средний и задний. Дальнейшая дифференцировка приводит к образованию пяти отделов головного мозга. Передняя часть переднего пузыря у большинства позвоночных, разрастаясь, дает собственно передний мозг (telencephalon), который у большинства позвоночных, разделяясь продольной складкой, образует обособленные полушария мозга; заключенные в них полости называются боковыми желудочками. Задняя часть переднего пузыря превращается в промежуточный мозг (diencephalon), образующий парные боковые выпячивания – глазные пузыри, из которых в дальнейшем формируется сетчатка и пигментная оболочка глаза; полость промежуточного мозга получает название третьего желудочка, а его полость называется сильвиевым водопроводом. Передняя часть заднего пузыря образует выступающий вверх задний мозг, или мозжечок (cerebellum).Остальная часть заднего пузыря разрастается в продолговатый мозг (myelencephalon), без резкой границы переходящий в спинной мозг. Полость продолговатого мозга – четвертый желудочек, или ромбовидная ямка, – продолжается в полость спинного мозга – невроцель. Эти пять отделов головного мозга характерны для всех позвоночных животных, но степень их развития, детали строения и функции существенно отличаются в разных классах (рис. 38).
Нервные клетки позвоночных животных, в общем сходны с нейронами других животных, но имеют свои особенности; их тела и короткие отростки – дендриты – составляют серое вещество мозга, а длинные отростки нервных клеток – аксоны, или невриты, окруженные неврилеммой (шванновскими клетками) и миелиновой оболочкой, образуют белое вещество. Оболочка изолирует нервные волокна, обеспечивая независимость проведения импульса (подобно изоляции электрических проводов). В организме позвоночного животного, где одновременно получается, передается и обрабатывается огромное количество разнообразной информации, подобное усложнение нервных путей необходимо (рис. 39).
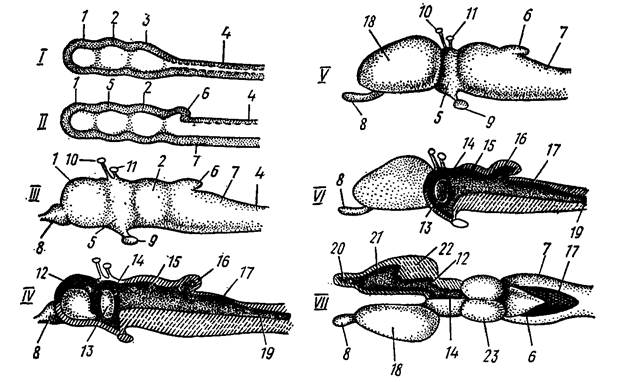

Рис. 38. Схема развития головного мозга позвоночных
(по Н. П. Наумову, Н. Н. Карташеву, 1979).
Основная масса серого вещества переднего мозга расположена в виде полосатых тел на дне и в его парных выпячиваниях – обонятельных долях. От каждой обонятельной доли отходят обонятельный тракт, образующий у обонятельной капсулы расширение – луковицу. Короткий обонятельныйнерв (nervus olfactorius) (I пара головных нервов) состоит из чувствующих волокон, связывающих клетки эпителия обонятельной капсулой с луковицей. Тонкий слой серого вещества выстилает стенки желудочков. Верх переднего мозга – мантия у большинства низших позвоночных образован только белым веществом; лишь у двоякодышащих рыб и земноводных в ее поверхностном слое появляется небольшое число нервных клеток. У пресмыкающихся в мантии имеется серое вещество (скопление нервных клеток), представляющее зачаток коры больших полушарий. У птиц объем переднего мозга резко возрастает, но сохраняется тот же тип строения, что и у пресмыкающихся. У млекопитающих величина переднего мозга увеличивается не столько за счет разрастания полосатых тел, сколько путем развития серого вещества на поверхности мантии.
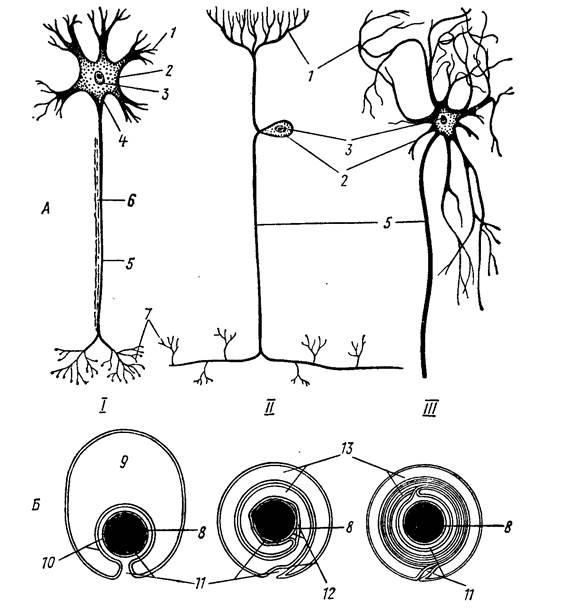

Рис. 39. Три типа нейронов (А) и этапы формирования оболочек аксона (Б): I – двигательный нейрон (мотонейрон), II – чувствительный (рецепторный) нейрон, III – нейрон центральной нервной системы (по Н. П. Наумову, Н. Н. Карташеву, 1979).
Этот слой нервных клеток, функционирующий как высший ассоциативный центр, называют корой больших полушарий. Его подразделяют на первичную кору, или гиппокамп (archipallium seu hipocampus) – разрастание образовавшегося у пресмыкающихся зачатка коры, и на вторичную кору (neopallium). На ней образуются борозды (извилины), за счет которых возрастает наружная поверхность коры.
Утолщенные стенки промежуточного мозга называют зрительным буграми (thalami optici); выросты их стенок образуют зрительные нервы (n. optici) (II пара головных нервов), имеющих хиазму (перекрест): часть волокон нерва правой стороны уходит в левый нерв, а часть волокон слева переходит в правый нерв. Зрительные нервы иннервируют только сетчатку глаза. Позади хиазмы на дне промежуточного мозга образуется полый вырост – воронка (infundibulum), к передней стенке которого примыкает железа внутренней секреции – гипофиз (hypophysis). На тонкой крыше промежуточного мозга развиваются два пузыревидных образования: передний называется теменным или париетальным органом, а задний – пинеальным органом или эпифизом (epyphysis). У круглоротых оба образования выполняют роль светочувствительных органов; оба несут и секреторную функцию. У остальных позвоночных животных эпифиз функционируй только как железа внутренней секреции. Теменной орган сохраняется у некоторых рыб, земноводных и у части пресмыкающихся как светочувствительный орган, у остальных позвоночных исчезает.
Крыша среднего мозга приподнята парными зрительными долями и образует двухолмие, у млекопитающих оно превращается в четверохолмие. Здесь в сером веществе заканчиваются волокна зрительного тракта. Начиная с двоякодышащих рыб и земноводных, значение среднего мозга как зрительного центра уменьшается. От него отходят две пары головных нервов, иннервирующих мышцы глаз: глазодвигательный нерв (n. oculomotorius) (III пара) и блоковой (n. trochlearis) (IV пара).
У круглоротых, части рыб и земноводных задний мозг имеет вид небольшой складки – мозжечка, ограничивающий спереди крышу четвертого желудочка. У хрящевых рыб он разрастается, прикрывая заднюю часть среднего и переднюю часть продолговатого мозга. У пресмыкающихся и, особенно у птиц и млекопитающих, размеры мозжечка еще более возрастают, а поверхность его коры, образованной серым веществом, увеличивается благодаря образованию глубоких и сложных складок. Мозжечок – центр координации движения и равновесия; участвует он и в регуляции тонуса мышц и других физиологических процессов.
Продолговатый мозг имеет утолщенное дно и стенки, тогда как крыша его объемистой полости – четвертого желудочка (ромбовидной ямки) – образована лишь тонким эпителием и сосудистым сплетением мягкой мозговой оболочки. Значительную массу продолговатого мозга составляет белое вещество (в том числе и волокна, приходящие сюда из спинного мозга); его разрастание по дну мозга образуют так называемые пирамиды. Серое вещество располагается в верхней части дна и стенок продольного мозга; от него отходят V-X (у млекопитающих – V-XII) пары головных нервов.
V пара – тройничный нерв (n. trigeminus) берет начало от боковой поверхности продолговатого мозга и тут же разделяется на три основные ветви: глазничную, верхнечелюстную и нижнечелюстную; иннервирует зубы и мускулатуру челюстной дуги, слизистую рта и кожу передней части головы. От дна мозга отходят тонкий отводящий нерв (n. abducens) (VI пара), иннервирующий наружную, прямую мышцу глаза. За тройничным нервом несколькими корешками, сливающимися в крупный ганглий, отходит VII пара – лицевой нерв (n.facialis). Он распадается на мощную глазничную (идет вместе с одноименной ветвью V нерва), щечную, небную и подъязычную ветви; лицевой нерв иннервирует кожу головы, слизистую ротовой полости и мускулатуру подъязычной дуги. Далее начинается короткий слуховой нерв (n. acusticus), VIII пара, иннервирующий чувствующий эпителий внутреннего уха. Языкоглоточный нерв (n. glossopharingeus), IX пара, отходит позади слухового нерва и иннервирует слизистую глотки и мускулатуру первой жаберной дуги, а у наземных позвоночных – глотку, ее мускулатуру и вкусовые тельца языка. От заднебоковой поверхности продолговатого мозга многочисленными корешками, сливающимися сразу в крупный ганглий, отходит Х пара – блуждающий нерв (n. vagus), разделяющийся затем на ряд ветвей: четыре жаберных (иннервируют слизистую глотки и мускулатуру II-V жаберных дут), внутренностную (иннервируют сердце, кишечник, плавательный пузырь, легкие), боковую (иннервируют органы боковой лини; редуцируется у наземных позвоночных). XI пара – добавочный нерв (n. accessorius) образуется из задних корешков блуждающего нерва; хорошо обособлен только у млекопитающих и слабо различим у других амниот; иннервирует мускулатуру плечевого пояса. XII пара – подъязычный нерв (n. hypoglossus) хорошо выражен только у амниот, отходит от брюшной стенки задней части, продолговатого мозга к мускулатуре языка и подъязычному аппарату. I, II, VIII пары головных нервов – только чувствующие, III, IV, VI, Xи XII – только двигательные, V, VII, IX и X – смешанные (включают чувствующие и двигательные волокна) (рис. 40).
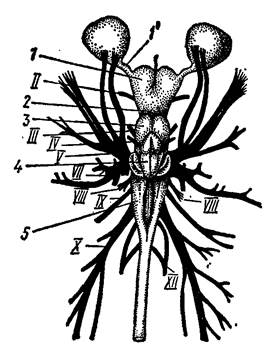

Рис. 40. Схема отхождения головных нервов у хрящевых рыб (по Н. П. Наумову, Н. Н. Карташеву, 1979).
Без отчетливых внешних границ продолговатый мозг переходит в спинной, который лежит в канале, образованном верхними дугами позвонков. Он имеет форму плоской ленты (крутлоротые) или округлого тяжа; в нем имеется небольшая полость – невроцель. Вокруг нее концентрируется серое вещество (скопление нервных клеток с их дендритами и опорных клеток); у наземных позвоночных на поперечном разрезе спинного мозга оно по своему очертанию напоминает крылья бабочки. В отличие от головного мозга в спинном мозге белое вещество (проводящие пути, образованные длинными отростками – аксонами нервных клеток с вкрапленными между ними опорными клетками) расположено снаружи. От боковой поверхности спинного мозга метамерно отходят спинномозговые нервы. От спинного выроста (рога) серого вещества отходит спинной корешок, состоящий из чувствующих волокон (передают нервные импульсы в головной мозг); на нем образуется расширение – спинномозговой узел (нервный ганглий). От брюшного выроста (рога) серого вещества отходит брюшной корешок, состоящий только из двигательных волокон (передают импульсы двигательной мускулатуре). Спинной и брюшной корешки сливаются в спинной нерв, выходящий между верхними дугами соседних позвонков и почти сразу же распадающийся на три ветви: спинную (иннервирует кожу и мышцы спины), брюшную (идет в боковые и брюшные стенки тела) и внутренностную (соединяется с ганглием симпатической нервной системы) и иннервирует внутренние органы. Каждая из них включает как двигательные (эфферентные), так и чувствующие (афферентные) волокна.
В спинном мозге замыкаются рефлекторные дуги, независимо от головного мозга обеспечивающие элементарные безусловно рефлекторные акты, как, например, оборонительные. Нейроны, связывающие правые и левые стороны каждого сегмента спинного мозга, увеличивают эти возможности. Однако работа всего спинного мозга подчинена контролю головного мозга, осуществляемого через так называемые спинальные тракты – совокупности нервных волокон, по которым передается информация в головной мозг (восходящие) или распространяются импульсы, идущие от головного мозга к сегментам спинного мозга (нисходящие пути). Существование параллельных иерархически организованных каналов связи – важное условие эффективного управления работой всех систем организма, обеспечивающее ее изменение соответственно внешним условиям (рис. 41).
Такая адаптивность увеличивается и вегетативной нервной системой, возникающей в результате разрастания участков ганглионарных пластинок. При этом по бокам позвоночного столба образуются цепочки ганглиев вегетативной симпатической и парасимпатической нервной системы. Длинные отростки клеток этих ганглиев направляются частично к внутренним органам (чувствующие и двигательные симпатические нервные волокна), частично входят в состав спинных корешков спинномозговых нервов. У двоякодышащих и костистых рыб и у всех наземных позвоночных ганглии вегетативной системы связаны друг с другом продольным нервным тяжем (truncus sympaticus).
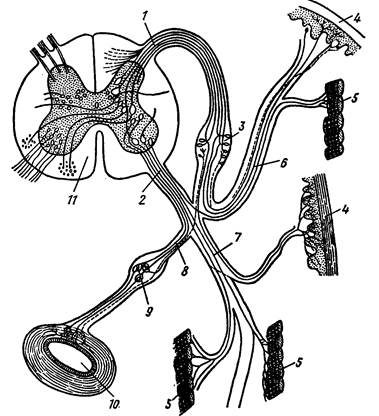

Рис. 41. Поперечный разрез спинного мозга (по Н. П. Наумову, Н. Н. Карташеву, 1979).
Соединительные ветви ганглиев обеспечивают связь центральной и вегетативной нервной системы. Чувствующие и двигательные волокна нервов, отходящих от симпатических ганглиев, иннервируют внутренние органы, не подчиняющиеся волевому контролю (сердце, кишечник, почки, половые и эндокринные железы и др). Показано, что вывод из строя вегетативной системы не ведет к гибели животного, но значительно снижает шансы выживания при отклонениях условий существования от нормы и при перенаселении.
Взаимосвязи центральной и вегетативной нервной системы создают возможность известной взаимозаменяемости нервных центров.
Гормональная система представлена разнообразными железами внутренней секреции. Ее сложность нарастает в эволюционном ряду позвоночных. При этом число и химизм участвующих в регуляции метаболизма гормонов в разных классах меняется мало, но существенно усложняется их функции – меняется реакция органов (тканей) на воздействие гормонов.
Деятельность отдельных звеньев гормональной системы взаимно координирована и находится под контролем центральной нервной системы; возникает сложный нейрогумолярный аппарат, координирующий все процессыв организме, включая особенности поведения и реакций на биотические факторы внешней среды. Развитие нейрогомулярного аппарата в ряду позвоночных животных приводит к становлению более сложных связей с окружающей средой и обеспечивает возрастание гомеостаза – внутренней устойчивости организма при меняющихся внешних воздействиях. Наконец, он участвует и в регуляции внутривидовых отношений и межвидовых связей в сообществе.
Органы чувств.Развитие и дифференцировка центральной нервной системы сопровождались возникновением и совершенствованием органов чувств – зрения, слуха и равновесия, обоняния, вкуса, восприятия движения воды (органы боковой линии водных позвоночных). Это улучшало возможности ориентации в пространстве, необходимость которого росла по мере увеличения подвижности животных.
Органы зренияпозвоночных животных представлены парными глазами более или менее шаровидной формы, лежащими в орбитах – впадинах черепа. Слой соединительной ткани или хряща образует наружную оболочку глаза – склеру. На передней поверхности глаза (перед зрачком) склера переходит в тонкую прозрачную роговицу, покрытую прозрачным эпителием. Изнутри к склере прилегает сосудистая оболочка, обильно снабженная кровеносными сосудами, за которой следует темная пигментная оболочка. На границе между склерой и роговицей края сосудистой и пигментной оболочек образуют кольцевую складку – радужину, ограничивающую отверстие – зрачок, который может расширяться и сужаться благодаря присутствию в радужине радиальных и кольцевых мышечных волокон. Насыщение радужины пигментами обеспечивает различную окраску глаза. Позади радужины образуется кольцевой валик – ресничное тело, представляющее скопление мышечных волокон, прикрепляющихся к оболочке хрусталика. Ресничное тело участвует в аккомодации глаза, изменяя форму хрусталика, и, у части наземных позвоночных, перемещая его по отношению к сетчатке. За зрачком лежит круглое или сплюснутое в двояковыпуклую линзу прозрачное тело – хрусталик.
Внутренний слой стенки глаза (сетчатка) тесно примыкает к пигментной оболочке. Она сложно устроена и состоит из светочувствительных (рецепторных), нервных и опорных клеток. К пигментному слою примыкает слой фоторецепторов, наружные членики которых имеют вид палочек или колбочек, и содержит светочувствительный пигмент. Ядра светочувствительных клеток образуют наружный клеточный слой, далее идет наружный сетчатый слой, где конечные разветвления чувствующих клеток контактируют с дендритами биполярных клеток. Отростки последних соприкасаются с дендритами ганглиозных клеток во внутреннем сетчатом слое. Ганглиозные клетки дают длинные нервные волокна (аксоны), образующие внутренний слой сетчатки и далее составляющие зрительный нерв. В месте выхода зрительного нерва светочувствительный слой сетчатки редуцируется, и этот участок называется "слепым пятном". В центре сетчатки обычно располагается область повышенной оптической чувствительности – желтое пятно; в его центре часто имеется углубление – ямка, где плотность фоторецепторов и нервных клеток увеличена. Внутренняя полость глазного бокала (яблока) заполнена прозрачным студенистым стекловидным телом.
Движение глаза в орбите обеспечивается сокращением четырех прямых и двух косых мышц глаза, прикрепляющихся к стенкам глазницы и к склере. Вокруг глаза развивается складка кожи, которая у наземных позвоночных, начиная с амфибий, превращается в подвижное верхнее и нижнее веки. У многих позвоночных развивается еще тонкое полупрозрачное третье веко – мигательная перепонка. В области век развиваются железы, секрет которых смачивает роговицу и предохраняет ее от высыхания.
Схема строения глаза едина во всех классах позвоночных. Отличия сводятся к частным деталям строения и связанных с ним образований – век, глазных желез. Уже указывалось на существование у круглоротых добавочных светочувствительных органов: париетального (теменного) и пинеального (эпифиз). У части рыб, некоторых амфибий и пресмыкающихся имеется теменной орган, принимающий глазоподобное строение: утолщенная передняя стенка образует хрусталик, задняя стенка пигментирована и содержит светочувствительные клетки, палочковидные окончания которых (в отличие от настоящего глаза) направлены к внутренней полости теменного органа; под светочувствительными клетками в слое нервных волокон лежат ганглиозные клетки, отростки которых образуют нерв, идущий в крышу промежуточного мозга (рис. 42).
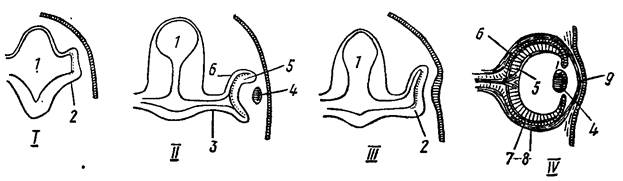
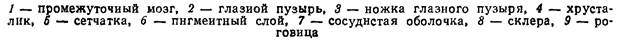
Рис. 42. Последовательные стадии развития глаза (по Н. П. Наумову, Н. Н. Карташеву, 1979).
Органы слуха анатомически связаны с органами равновесия; у позвоночных животных они всегда парные (правое и левое "ухо").
У водных позвоночных такой комплексный орган образован капсулами внутреннего уха, закладывающимися по бокам головы зародыша в виде парных утолщений эктодермы. Каждая из них превращается сначала в ямку, а затем в пузырек, сообщающийся с поверхностью каналом, и позднее отшнуровывающийся от эктодермы. Слуховой пузырек перехватом разделяется на два отдела: верхний и нижний. Из верхнего формируется овальный мешочек и соединенные с ним три полукружные каналы, лежащие в трех взаимно перпендикулярных плоскостях, – орган равновесия (вестибулярный аппарат); нижний отдел превращается в круглый мешочек, от которого отходят полый выступ – лагена, у высших позвоночных образующий спирально завитой канал (улитку) – орган слуха.
Эпителий, выстилающий перепончатый лабиринт, на некоторых участках кругловатого и овального мешочка и в полукружных каналах имеет чувствующие клетки, снабженные упругими волосками, вдающимися в просвет внутренней полости; основания грушевидных чувствующие клеток оплетаются разветвлениями конца слухового нерва. Внутренняя полость перепончатого лабиринта заполнена эндолимфой, в которой взвешены мелкие кристаллики извести – отоконии, иногда сливающиеся в,крупные образования (отолиты костистых рыб). Всякое изменение положения головы вызывает перемещение эндолимфы и отокониев, раздражающих при этом чувствующие клетки. Перепончатый лабиринт, таким образом, функционирует как орган равновесия. Звуковые волны, вызывающие колебание эндолимфы и находящихся в ней включений, также вызывают раздражение чувствующих клеток. Однако слуховую функцию у рыб несут чувствующие поля полого выроста круглого мешочка. У наземных позвоночных его размеры заметно увеличиваются, и в образовавшейся таким путем улитке происходит усложнение воспринимающего аппарата за счет разрастания чувствующих клеток и добавочных мембран. Наряду с усложнением воспринимающего аппарата внутреннего уха у наземных позвоночных образуется среднее ухо, снабженное барабанной перепонкой и слуховыми косточками, а затем формируется и наружное ухо, хорошо развито у млекопитающих). Эти преобразования обеспечивают возможность слуха в воздушной среде, отличающейся более слабой звукопроводимостью. Часть лабиринта, функционирующая в качестве органа равновесия (овальный мешочек, полукружные каналы), у всех позвоночных сохраняется примерно на одном уровне развития; лишь у круглоротых он видимо, вторично упрощен (рис. 43).
У водных позвоночных имеются специальные сейсмосенсорные кожные органы боковой линии. Они служат преимущественно для восприятия слабых колебаний и токов воды. У круглоротых и живущих в воде амфибий эти органы представляют собой лежащие на поверхности или в мелких ямках утолщения небольших участков эктодермы, в которых среди опорных клеток лежат грушевидные чувствующие клетки. Их выдающиеся над поверхностью части снабжены щетинками (жгутиками), а основания оплетены концевыми окончаниями боковой ветви блуждающего нерва. Эти органы располагаются в 1-3 продольных ряда на боковой поверхности тела, иногда в виде сложной сети, на голове. У рыб в толще кожи образуются каналы, открывающиеся наружу специальными отверстиями; у костистых рыб эти отверстия пронизывают чешуи и покровные кости. На стенках каналов располагаются отдельные рецепторы боковой линии, имеющие описанное строение.
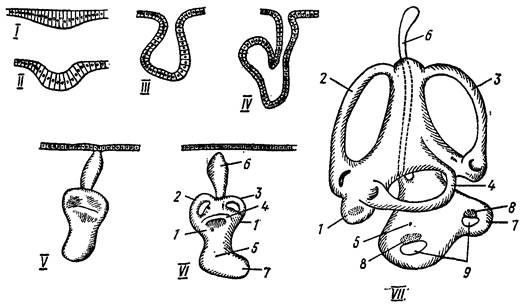

Рис. 43. Схема развития внутреннего уха позвоночных животных: I-VII – последовательные стадии (по Н. П. Наумову, Н. Н. Карташеву, 1979).
У рыб встречаются и отдельные органы боковой линии, лежащие на поверхности кожи. Улавливая органами боковой линии слабые токи воды, водные позвоночные могут ориентироваться в течениях, а воспринимая волны, идущие от плывущих животных или отраженные волны собственного перемещения, могут обнаруживать приближение хищника или добычи, плыть, не натыкаясь в темноте на подводные предметы. У наземных позвоночных органы боковой линии исчезают (рис. 44).
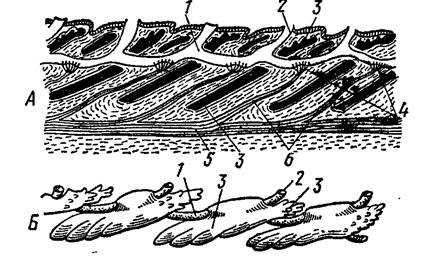

Рис. 44. Схема органа боковой линии костистой рыбы: А – продольный разрез, Б – вид сбоку (по Н. П. Наумову, Н. Н. Карташеву, 1979).
Орган обоняния(рис. 45)развивается как утолщение эктодермы, которое затем погружается в кожу, образуя обонятельный мешок, открывающийся наружу отверстием – ноздрей. Эпителий обонятельного мешка состоит из опорных и удлиненных чувствующих клеток. Выходящие на поверхность эпителия концы чувствующих клеток несут пучок волосков, а проксимальные концы их соединяются с волокнами обонятельного нерва. У круглоротых парные вначале обонятельные мешки срастаются в один мешок, открывающийся наружу одной ноздрей. У остальных позвоночных органы обоняния парные. У некоторых рыб каждая ноздря разделяется кожистой перемычкой на два отверстия – через одно вода поступает в обонятельный мешок, а через второе выводится наружу. Образование складок на стенках обонятельного мешка увеличивает поверхность чувствующего эпителия. У двоякодышащих и кистеперых рыб полость обонятельного мешка через ноздрю открывается наружу, а через внутреннюю ноздрю хоану сообщается с ротовой полостью.
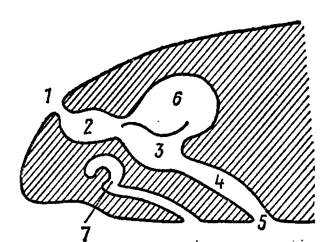

Рис. 45. Дифференцировка обонятельного мешка у ящериц (разрез) (по Н. П. Наумову, Н. Н. Карташеву, 1979).
Переход к воздушному дыханию и выход на сушу сопровождался превращением обонятельного тракта в дыхательно-обонятельный. Воздух через наружные ноздри проходит в обонятельный мешок и через хоаны поступает в ротовую полость, гортань и легкие. В связи с этим уже у aмфибий обонятельный мешок начинает подразделяться на дыхательный или респираторный (выстлан простым эпителием), и обонятельный, или ольфакторный (выстлан чувствующим эпителием) отделы. В обонятельном отделе обычно появляются складки, увеличивающие его поверхности. Еще отчетливее дифференцировка у настоящих наземных позвоночных. Особенно сложно строение обонятельного отдела у млекопитающих, у которых развиваются ажурные переплетения решетчатой кости, образующей костные обонятельные раковины с очень большой общей поверхностью, выстланной обонятельным эпителием.
Органами вкуса у позвоночных служат вкусовые почки, представляющие скопление чувствующих и опорных клеток. Основания чувствующих клеток оплетаются концевыми окончаниями нервов (лицевого, а у млекопитающих еще и языкоглоточного). Вкусовые почки у рыб есть впищеводе, глотке и ротовой полости, а также на губах, усиках, иногда на плавниках и других участках тела, у остальных позвоночных они сосредоточены в ротовой области (преимущественно на языке и иногда в начальной части глотки).
Восприятие тактильных, химических, электрических (электромагнитных), температурных и других раздражений обеспечивается наличием свободных окончаний чувствующих нервов по всей поверхности кожи. В некоторых случаях они представляют специальные органы (электрорецепторы хрящевых рыб) или входят в состав системы органов боковой линии. Простое строение (свободные нервные окончания) имеют и так называемые интерорецепторы, собирающие информацию о состоянии внутренне среды и органов тела животного – химизме, температуре, осмотическом состоянии и других показателях. Информация от органов чувств (экстерорецепция) и интерорецепторов поступает в соответственные отделы головного мозга, регулирующие физиологические параметры и поведение животного.
Итак, нервная система возникла путем обособления чувствующих эктодермальных клеток, которые погружались глубже под защиту поверхностного эпителия. У предков хордовых животных, вероятно, имелась продольная спинная полоса чувствующего эпителия, которая вся целиком углубилась сначала в виде открытого желоба, а затем образовала замкнутую трубку. Аналогичное явление наблюдается на органах боковой линии позвоночных. Эти органы также погружаются сначала в открытую борозду, которая затем замыкается в канал. Что нервная трубка хордовых животных первоначально имела значение органа чувств, видно из того, что у низших хордовых последние находятся еще в самом мозге. У ланцетника весь мозг является светочувствительным, так как внутри его стенок имеются особые чувствующие клетки, частью окруженные пигментными клетками и образующие так называемые глазки Гессе. Обонятельная ямка ланцетника развивается в связи с невропором. У позвоночных животных органы зрения развиваются всегда за счет самого мозга, а орган обоняния по своему происхождению связан с невропором, развивается за счет эктодермального утолщения, непосредственно прилежащего к переднему концу нервной пластинки. Наконец, органы боковой линии и органы слуха, так же как и нервные узлы, развиваются за счет эктодермы. Эпителий, выстилающий центральный канал нервной трубки, первоначально является мерцательным и вызывает движениями своих ресничек ток воды, входящий через невропор и направленный назад через нервно-кишечный канал наружу. Это обстоятельство освещает значение этих отверстий. Как и раньше, на стадии нервной пластинки и открытого желоба, мерцательные реснички производят постоянный обмен воды, что было необходимо, если этот орган служил для химического испытания (обоняние, вкус). Это объясняет и возникновение органа обоняния в связи с невропором. Наконец, постоянный обмен воды мог иметь огромное значение в смысле осуществления более интенсивного газового обмена (дыхания) нервной ткани, что было чрезвычайно важно при слабой дифференцировке органов дыхания и кровообращений.
Таким образом, центральная нервная система хордовых животных непосредственно с центральными органами беспозвоночных не сравнима, концентрация элементов субэпителиальной нервной сети, мозг хордовых развивался путем обособления от эктодермы особой чувствующей пластинки.
Дата добавления: 2022-04-12; просмотров: 288;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
