ПАЙДА БОЛУ МЕХАНИЗМІ. 10 глава
D = D1 + D2 + D3 + …
Линза арқылы алынған кескіндегі кемшіліктерді аберрация деп атайды. Аберрация латын сөзі (aberratio), ол ауытқу дегенді білдіреді. Егер аберрация күрделі жарық (монохроматы емес) кезінде байқалса, оны хроматты аберрация деп, басқа кездегілері монохроматты аберрация деп аталады, өйткені мұндай ауытқулардың пайда болуы жарықтың спектріне байланысты емес. Осындай ауытқулардың кейбір түрлерін қарастырайық.
1. Сфералық аберрация деп, өте жұқа болып келетін линзаның шеткі аймағы арқылы өткен жарық ағыны (1), оның орта бөлігі арқылы өткен жарықтан (2) көбірек сынады (ауытқиды), нәтижесінде линзаның шеті мен орта бөлігі арқылы өткен жарық ағыны экранда бір біріне сәйкес келмейтін екі түрлі кескін жасайды (4-сурет).

4- сурет
Мұндай ауытқуларды болдырмау үшін жинағыш және шашыратқыш линзалар жүйесін бірге қолданады.
1. Астигматизм деп, жарық ағыны линза осіне үлкен бұрышпен, қиғаштап түсу нәтижесінде пайда болатын ауытқуды атаймыз. Мысалы, сәуле линзаға белгі бір бұрышпен түскен кездегі нүктенің кескіні элипс түріндегі дақ болса, басқа түсу бұрышында сол нүктенің кескіні түзу сызық болады.
Көзде мұндай құбылыс көз бұршағы мен мүйізді қабат беттерінің қисықтығында асимметрия болса байқалады, оны болдырмау үшін, қистықтық радиусы түрлі бағытта әр түрлі болып келетін линза қолданылады.
2. Дисторсия, деп линзаға түскен сәуле бағыты мен линзаның оптикалық осі арасындағы түсу бұрышы үлкен болғанда пайда болатын ауытқуды атайды. Бұл кезде, пайда болған дене кескіні оның шын мәнідегі түрінен өзгеше болады, мысалы, квадратты тордың кескіні «жастық»(а) немесе «бөшке»(б) тәрізді түрде көрінеді (5-сурет). Мұндай ауытқуды болдырмау үшін бір-біріне кері дисторсиялық қасиеті бар линзалар жүйесін құрайды.

a) б)
5 сурет
3. Хроматтық аберрация деп, линза арқылы алынған дене кескіні айналасының түрлі түсті болуын атаймыз. Мұндай құбылыс ақ жарық құрамындағы сәулелердің сыну көрсеткіштерінің олардың толқын ұзындығына байланысты болуынан. Мысалы, күлгін сәуле қызылға қарағанда көп сынады, сондықтан әр түс өз кескінін жасайды, нәтижесінде экранда пайда болған кескіннің төңірегінде түрлі түс пайда болады.

6-сурет
Көз кез келген қашықтықта жатқан нәрсені анық көруі тиіс. Енді осы құбылысты қарастырайық. Жұқа линзаның формуласы мына түрде жазылады:
1/d + 1/f = 1/F, (1)
мұндағы d- нәрседен көзге дейінгі қашықтық, f- линзадан кескінге дейінгі қашықтық, F- линзаның фокустық қашықтығы. Бұл теңдеудегі f көз бұршағынан тор қабатқа дейінгі қашықтық, ол тұрақты шама, ал d үздіксіз өзгеріп отыратын шама. Егер нәрседен көзге дейінгі қашықтық үнемі өзгеріп отырса, онда 1-теңдік орындалуы үшін теңдеудегі F үнемі өзгеріп отыру тиіс. Оған көз бұршағы бетінің қисықтығын өзгерту арқылы қол жеткіземіз. Бұл құбылыс былайша жүреді: көз бір нәрседен қашықтығы өзгеше екінші нәрсеге ауысқанда, екінші дененің кескіні анық көрінбейді, (өйткені d өзгерді, ал F сол қалпында) бұл туралы миға тиісті сигнал жетеді, мидан шыққан кері сигнал көздегі циллиарлы бұлшық етің жиырылуын немесе созылуын тудырады, соның нәтижесінде көз бұршағының фокусы дене кескіні анық болғанға дейін өзгереді, әрине бұл үрдіс өте жылдам жүреді көздің мұндай қабілетін аккомодация деп атайды.
Көзі қалыпты көретін адамның көзі дене кескінін көздің тор қабатына фокустайды. Кейбір адамдардың көздері кескінді тор қабаттың алдына, оған жеткізбей түсіреді (7а- сурет), көздің мұндай кемшілігін жақыннан көргіштік деп атайды. Мұндай адамдар жақын жатқан нәрселерді анық көргенімен, алыс жатқан денелерді бұлыңғыр көреді, Мұндай көз ақауын түзету үшін шашыратқыш линзадан жасалған көзілдірік киеді.
Кейде адам көзі дене кескінін тор қабаттың сыртына фокустайды (7б- сурет), көздің мұндай кемшілігін алыстан көргіштік деп атайды. Мұндай адамдар алыс жатқан денелерді анық көргенімен, жақын жатқан нәрселерді бұлыңғыр көреді, Мұндай көз ақауын түзету үшін жинағыш линзадан жасалған көзілдірік киеді.
| |
| |
|
|
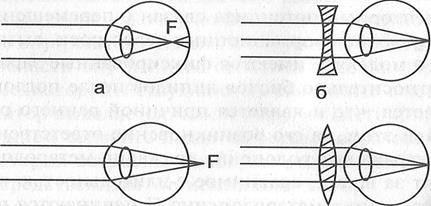
7 –сурет
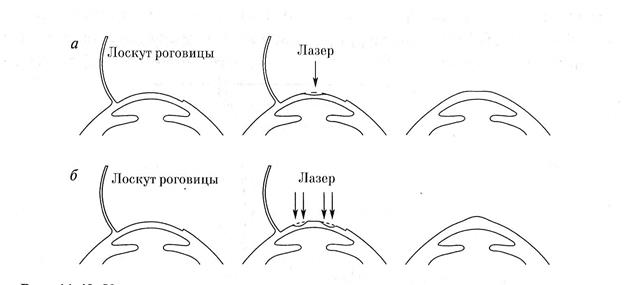
Қазірге уақытта мұндай көздің кемшіліктерін мүйіз қабаттың алдыңғы бетінің қисықтығын өзгерту арқылы дұрыстайды. Оның бірнеше әдістермен іске асырады. Соның бірі радиалды кератотомия (РТ) деп аталды және ол мына түрде орындалады, мүйізді қабаттың орта бөлігін радиал бойымын төрт немесе сегіз сызық түрінде сызады(кеседі), осылайша мүйізді қабат қалыңдайды, нәтижесінде оның оптикалық күші кемиді, бұл өз кезегінде көз миопиясын өзгертуге алып келеді (8-сурет).

8 сурет
Келесі әдіс фоторефракциялыкератотомия (ФРК) немесе лазерлы кератомилеза in situ(LASIK) деп аталады, мұндай ота мүйізді қабаттың қисықтығын азайтады (миопия болса) немесе жоғарылатады (гиперопия болса). ФРК немесе LASIKәдістері лазер сәулесімен мүйізді қабатың орта бөлігін(стома) қыздыру арқылы оның булануына қол жеткіземіз, осы арқылы мүйізді қабаттың қисықтығын өзгертеміз.
Бұл былайша іске асырлады, ФРК кезінде мүйізді қабаттың сырты беті, эпителий қабаты лазер сәулесі арқылы жұқартылады, ал LASIK кезінде мүйізді қабаттың сыртқы бетіннен қалыңдығы 20% болатындай қабатты ашып (сылып), ашық қалған мүйізді қабаттың бетіне лазер сәулесімен әсер етіп, оны жұқартамыз, нәтижесінде оның қисықтығы өзгереді, онан соң мүйізді қабатты сылынған өте жұқа қабатпен қайта жабамыз
Енді көрудің биофизикалық негізін талдайық. Жоғарыда айтқандай, көзге түскен жарық ағыны көз бұршағы арқылы жарықты сезгіш жасушаларға-таяқшалар мен құтышаларға фокусталады. Таяқшалар тор қабаттың сфера бетін жартылай алып жатыр және ол кескінді «ақ – қара» түрінде көруге мүмкіндік береді. Құтышалардың саны аз, олар негізінен тор қабаттың орта бөлігін алып жатыр және ол кескінді «түрлі-түсті» түрінде көруге мүмкіндік береді. Алдымен көздің тор қабатының құрылысын талдайық, өйткені көру процесі негізіннен осы аймақта өтеді.
Көз торы бірнеше қабаттардан тұрады. Жарық түсетін бірінші қабат- нерв талшықтарынан (жасушаларынан) тұрады, олар таяқша мен құтышыларда пайда болған электр сигналдарын миға жеткізеді. Онан соң бірнеше қабаттар орналасқан, олардың негізгі қызметтерін адам физиологиясы курсында талдайтындықтан қазір қарастырмаймыз. Бұл қабаттардан кейін жарықты сезгіш рецепторлар - таяқша мен құтыша жасушаларынан тұратын қабат орналасқан. Олардан соң қара түсті пигменті меланин қабаты бар(9- сурет).



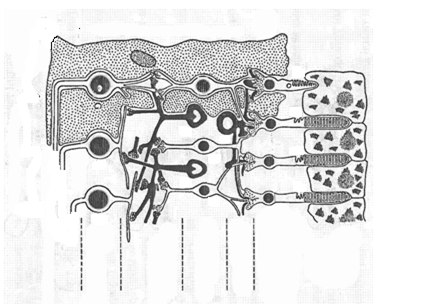
9-сурет. Көз торының құрылысы. 1-фоторецепторларда (таяқша мен құтышаларда) пайда болған электр сигналын миға жеткізетн нерв талшықтары. 2-ганглиоз жасушаларының қабаты. 3-ішкі синап қабаты. 4- ішкі ядролық қабат. 5-сыртқы синап қабаты.6- сыртқы ядролық қабат. 7- құтышалар мен таяқшалар қабаты. 8- эпителии (қара түсті пигмент қабаты).
Бір қарағанда таяқша мен құтыша жасушалары, көзге түсетін жарық ағыны бірден түсетін алдыңғы қатарда орналасуы қажет секілді, ал шындығында олар көз торының түбінде, ең соңғы қабатында жатыр.
Фоторецепторлардың мұндай орналасуы кездейсоқ емес, тор қабаттан өткен, көздің ішкі құрылымынан шағылған немесе шашыраған сәулелердің фоторецепторларға түсуіне жол бермейді, ондай сәулерді қара түсті меланин қабаты жұтады, осы арқылы көз таяқшалары мен құтышаларының сезгіштік қабілетін үнемі жоғары деңгейде болуына мүмкіндік береді және өте сезімтал болатын таяқша мен құтышаларды кездейсоқ сыртқы әсерлерден(сәулелерден) сақтайды.
Жарық ағыны сезгіш рецертораларға жеткенше мүйізді қабаттан, көз бұршағынан, шыны тәрізді дене және тағы да бірнеше қабаттардан өтуіне тура келеді және кей қабаттардан жарық шағылысады, кей қабатарда жұтылады, осылайша өзінің бастапқы энергиясын көп бөлігін жоғалтады. Көзге түскен жарық ағынының 4 % мүйізді қабатан шағылысады, ал 50% көз ішіндегі орталарда (көз бұршағында, шыны тәрізді ортада және т.б. қабаттарда) жұтылады, жарық энергиясының 40% көз торы қабаттары арқылы өтіп, ең соңғы қара түсті пигменті эпителии қабатында жұтылады. Осылайша жарық энергиясы бірте-бірте азайып, көз фоторецепторларына тек бірнеше пайызы ғана жетеді.
Соңғы кезде жүрізілген зерттеулер, жарықты сезгіш фоторецепторлардың күшейту коэффициенті 105-106 болатын трансформатор тәрізді энергиясы өте әлсіз жарықты көру сезімін тудыратын деңгейге жететіндей күшейтіп, онан соң электр сигналына айналдыратындығын көрсетті. Қараңғы жағдайға бейімделген көздің 491 нм толқынға сәйкес келетін 2-3 фотонын сезетіндігі анықталды, олай болса көзді сезімталдығы өте жоғары оптикалық құралдардар қатарына жатқызуға болады.
Қараңғыға бейімделген көздің ең төменгі жарықтылықты сезінуін абсолютік көру табалдырығы деп атайды, ол шамамен 2,1-5,7×10-17Дж ,бұл көк-жасыл түстерге сәйкес келетін 58-148 кванттың энергиясына тең.
Жарық ағыныңың интенсивтілігі мен оны көздің сезу арасындағы байланыс Вебер-Фехнер заңымен сипаталады және ол мына түрде беріледі:
Y =k×ln 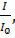
мұндағы I-жарық интенсивтілігі, I0 – көздің жарықты абсолютті сезу шегі, k- тұрақты шама.
Бұл заңдылықтан мынадай қорытынды жасауға болады: егер жарық интенсивтілігі логарифимді түрде өссе оны көздің сезуі сызықты түрде өзгереді.
Енді таяқша мен құтышаның құрылысымен танысайық. Көзде жалпы саны 125 млн таяқша мен 6,5 млн құтыша бар. Адам көзіндегі таяқшаларының ұзындығы 50-60 мкм, диаметрі 2 мкм болса, құтышалардың ұзындығы 30 мкм, диаметрі 6 мкм тең. Көз торындағы таяқша мен құтышаларға нерв талшықтары жалғанған, миллиондаған тармақтардан тұратын осындай талшықтар жинала келе көлденең қимасы 4 мм2 болатын көру нервіне айналады.
Көз таяқшалары өз ара байланысқан сыртқы және ішкі екі бөліктен(сегменттен) тұрады. Сыртқы бөліктің(сегмент) ұзындығы 20-30 нм, яғни таяқша ұзындығының жартысына тең. Сыртқы бөліктегі(сегмент) цитоплазма бір-бірінен ара қашықтығы 30нм болатын, жалпы саны 700 ден 1000 дейін жететін (сиырда-200, бақада-1500), сыртқы пішіні орта бөлігінен қысылған ауа шарына ұқсас, жарықты сезгіш дискілермен толтырылған. Дискі санының мұндай көп болуы фоторецепторлар алып жатқан беттің үлкен болуына алып келеді, бұл көздің сезгіштігін артырады. Әр дискінің диаметрі 2 мкм, қалыңдығы 15-18 нм тең. Дискі қабырғасының құрылысы биологиялық мембранадай, фосфолипид молекуларынан құралған. Фосфолипид қабатында родопсин молекулалары бар, оның кейбірі қабатқа жабысып, кейбірі оны тесіп орналасқан(10- сурет).
Көздің көруі, таяқша дискілеріне түскен жарық әсерінен жүретін бірінші реттік фотоқұбылыстарға байланысты болғандықтан, дискідегі мембраналарды фоторецепторлы мембраналар деп атайды.
Барлық биомембраналарға тән фоторецептордың мембранасы да фосфолипид және ақуыздан құралған, бірақ олардың бірнеше ерекшеліктері бар. Дискі мембранасындағы ақуыздың 90% көру пигменті - хромогликопротеид родопсин молекуласына тиісті.




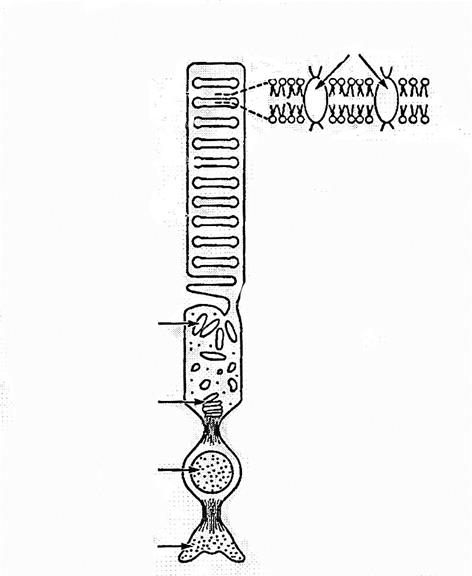
10- сурет. Көз таяқшасының құрылысы.
Мембранадағы фосполипид молекуласы құйрығының жоғары деңгейлі қанықпаған майқышқылынан тұруы(құралуы), биомембрананың гидрофобты құрылымының қозғалғыштық қасиетін жоғарылатып, оның тұтқырлығын 30 мПа×с дейін төмендетеді.
Биомембранадағы липид молекулаларының 80% астамының құрамына қанықпаған майқышқылы кіреді, бірақ холестерин және гликолипид молекулалары өте аз мөлшерде. Сондықтан дискі мембранасындағы бір родопсин молекуласына 90 фосфолипид және 6 холестерин молекуласы сәйкес келеді. Фоторецепторлы мембрана бетінің 75% үлесін липидтер, 25% үлесін родопсин алып жатыр. Дискі мембрана массасың 40-50% үлесі липидтерге, қалғаны родопсинге сәйкес келеді.
Фоторецепторлар үнемі жаңарып, ескі дискінің орнына жаңа дискі пайда болып отырады. Дискінің жаңаруы деп жұтылған жарық әсерінен родоспин молекуласының түссізденуін, яғни жарық квантарын сезбеуін және мембранадағы фосфолипидтардың жұтылған жарық әсерінен тотығуын (фотототығу феномені) атаймыз. Бұл құбылыс тәуліктікке және уақыт мерзіміне тәуелді. Мысалы, сыртқы бөліктегі таяқша ұштарындағы дискілер таңертең және күндіз жаңарса, сыртқы бөліктегі құтышалардағы дискілер кешкі және түнгі уақыт мезгілінде жаңарады. Ескі дискілер бөліктің ішімен қозғалып, эпителии пигментінің жасушаларында жұтылады. Мысалы, бір тәулік ішінде маймыл көзінің бір таяқшадасындағы 80-90 дискісі жаңараса, онда сыртқы бөлік ішіндегі барлық дискілер 9 -12 тәулік ішінде толығымен жаңарады және бұл құбылыс өмір бойы үздіксіз жүріп жатады.
Фоторецепторлардың тағы бір ерекшелігін қарастырайық. Олар өз ось бойымен, мембрана жазықтығына перпендикуляр бағытта жылдам айналады, сонымен қатар олардағы латериалдық диффузия деңгейі өте жоғары. Мысалы, родопсин үшін латериалды диффузия коэффициенті (3,5±1,5)×10-9 см2/с тең, сондықтан мембрана қабатындағы әр родопсин молекуласы бір секунд ішінде басқа молекулармен 105-106 рет соқтығысады.
Енді жарықты сезгіш пигментерді талдайық, өйткені олар көру құбылысының негізін қалайды. Көру дискілерінде 4 түрлі жарықты сезгіш пигменттің бар екендігі анықталды. Таяқшаларда мұндай пигментер - родопсин (адамдар мен теңіз жануарларында) және порфиропсин (тұщы су жәндіктерінде) деп аталса, құтышаларда- йодопсин мен цианопсин деп аталады. Шын мәнінде родопсин молекулалық массасы 40 000 тең, қызыл түсті, түскен жарық әсерінен ыдырайтын (түссізденетін), ақуыз молекуласы. Ал йодопсин болса, күлгін түсті, бірақ бұл молекуланың жарық әсерінен ыдырау жылдамдығы родопсинге салыстырғанда төмен, сондықтан оның жарықты сезу деңгей де төмен болады.
Родопсин молекуласы көрінетін жарық толқындарын жақсы жұтады, бұл толқындардың ішінен максимал түрде жұтылатыны lmax = 498 нм сәйкес келеді, сонымен қатар ол lmax= 278 нм сәйкес келетін ультракүлгін сәулесін де жұтады, бірақ оны жұтатын родопсин құрамындағы ретиналь емес, опсин құрамына енетін ароматты аминқышқылдарға жататын триптофан мен тирозин (11-сурет).


11 сурет
Ультракүлгін толқыны көздің түрлі қабаттарында толығымен жұтылу себепті родопсинды қоздыра алмайды, яғни көру сезімін тудырмайды. Егер ультракүлгін сәулесінің квантарының энергиясы жоғары болса, ондай сәуле көз құрылымын зақымдайды.
Родопсин жарықты сезгіш хромопротеидтерге жатады және ақуыздың бір түрі болып табылатын, өз ара байланысқан опсин мен ретиналдан құралған кешен. Шын мәнінде ретиналь альдегид тобына жататын, жарықты сезгіш- хромофор болып табылады. Ретиналь екі түрлі изомериялық пішінде болуы мүмкін, қарыңғы кезде, ол опсинмен бірігіп 11-цис-ретиналь деген түрде болса, родопсиннің жарық квантын жұтуы нәтижесінде ол опсинен ажырып, транс-ретиналь деген түрге ауысады, мұндай құбылыс фотоизомеризация деп аталады. Осының нәтижесінде (жұтылған жарық әсерінен) родопсин түссізденеді, мұны фотолиз құбылысы деп атайды.
Жұтылған жарық әсерінен ыдыраған родоспин қайта қалпына келеді және бұл құбылыс үздіксіз жүріп жатады. Арнаулы ретинальизомеразы ферменті арқылы транс-ретиналь қайтадан 11 -цис- ретинальға айналады және ол опсинге қосылып жаңа родопсинді түзейді. Егер көз торындағы жарық ағыныны тұрақты болса, онда родопсиннің ыдырауы мен оның қайта түзелуі тұрақты сипатта болады.
1967 ж. родопсиннің түрленуін алғаш зерттеген Д.Уолдқа Нобель сыйлығы берілді. Ол мұндай түрленудің бірнеше этаптан тұратындығын анықтады, алдымен родопсин жарық квантын жұтып t < 6×10-12c кейін батородопсинге айналады, ол t »3×10-8c кейін люмиродопсинге айналады, осылайша бірнеше түрленуден соң транс-ретиналь пайда болады. Бұл түрлену төмендегі схемада берілген және түрленудің тек бірінші этапы ғана жарық кезеңінде жүрсе, қалғандары қараңғы кезеңде жүреді (12-сурет).

12-сурет
12-сурет
Д.Уолд өз зерттеуінде родопсинді қараңғы жағдайда бөліп алып, оны температурасы -1400С болатын сұйық азотта ұстаған, осы күйдегі родопсинге жарық түсіргенде және оның температурасын жоғарылатқанда, родопсиннің жұтылу спектрі өзгеріп отырған, бұл родопсиннің тізбекті түрде, бір түрден екінші түрге ауысқандығын, яғни түрленгендігін көрсетеді
Әр кезеңдегі конформер (батородопсин, люмиродопсин, метародопсин және т.б.) транс-ретиналь күйдегі ретиналь болғандықтан, оған жарық квантымен әсер етсек, ол бірден родопсинге айналады (арнаулы ферментсіз).
Осындай фотоизомериялық өзгерістің нәтижесінде таяқшадағы тыныштық потенциалы өзгеріп, әсер потенциалы пайда болады. Көз таяқшасындағы осындай құбылыстар тізбегі көру сезімін тудырады.
Шындығында, фоторецепторлық жасушалардағы мембраналық потенциалдың өзгерісі таяқшаның сыртқы бөлігінің (сегмент) плазмолеммасында орын алады, ал родопсиннің фотоизомеризациясы таяқшаның сыртқы бөлігінің (сегмент) дискі мембранасында байқалады. Мембрана қасиетінің өзгеруі, оның құрамындағы родопсиндегі өзгерістермен (ретиналдың, опсиннің өзгеруі) қабаттасып жүреді.
Енді осы құбылыстарды талдайық. Алдымен мына ерекшелікпен танысайық, фоторецепторлардағы тыныштық потенциалының пайда болуы механизмінде біраз өзгешеліктер бар. Таяқша жарық квантарын жұтпаған кезде, яғни қараңғылық кезеңінде, тыныштық потенциалы таяқшаның ішкі бөлігінде (сегмент) плазмолеммада К+ иондарының әсеріннен пайда болса, таяқшаның сыртқы бөлігіндегі(сегмент) потенциал Na+ иондарының әсерінен туындайды, яғни таяқшадағы тыныштық потенциалы осы екі түрлі иондардың әсерінен пайда болады деп саналады.
Көз таяқшасының сыртқы бөлігіндегі(сегмент) тыныштық потенциалының таңбасы өзгеше, ішкі орта «оң», ал сырты «теріс», ал таяқшаның ішкі бөлігінде(сегмент) керісінше, ішкі орта «теріс», ал сырты «оң» зарядталған. Бұл құбылыс, таяқшаның сыртқы бөлігінің (сегмент) Na+ иондарын өткізетін каналдарының циклды гуанозин монофосфатты(цГМФ) әсерінен ашық болуынан екендігі анықталды. Сондықтан тыныштық күйдегі мембрана қабатының натрий иондарын өткізу коэффициенті басқа иондарға қарағанда басым болады, осының әсерінен және электрохимиялық градиент есебінен сыртқы ортадағы натрий иондары ішке қарай диффузияланады (пассивті тасымал). Осылайша, көз таяқшасының сыртқы бөлігінің(сегмент) цитоплазмасында жиналған натрий иондары, екі бөлімді қосатын орта арқылы ішкі бөлімге тасымалданады. Жиналған натрий иондары, митохондрия жеткізген АТФ энергиясының есебінен сыртқы ортаға шығарылады (активті тасымалдау). Осылайша, Na+ ионының ішке, ал К+ ионының сыртқа қарай қозғалысы нәтижесінде таяқшада тыныштық потенциалы орнайды (13 сурет).
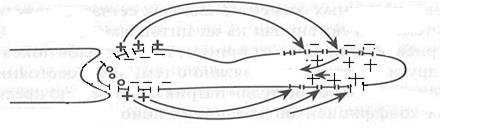
13 сурет. Таяқшаға жарық әсер етпеген кездегі тыныштық потециалының пайда болуы.
Дата добавления: 2020-11-18; просмотров: 821;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
