СЕЛЕКЦИЯ И РАЗВЕДЕНИЕ. Генетика собак
Как же узнать степень гомозиготности особи и не поможет ли нам разобраться с ней пресловутый коэффициент инбридинга?
В 20-х годах минувшего столетия известный американский генетик Сьюэл Райт вывел математическую формулу, с помощью которой можно рассчитать степень увеличения гомозиготности в популяции под влиянием близкородственного спаривания. Позже русский генетик Д. А. Кисловский слегка подкорректировал формулу, и теперь мы знаем ее как формулу Райта – Кисловского:
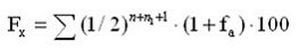
где Fx коэффициент инбридинга пробанда, %;
n, ni — ряды родословной;
fa — коэффициент инбридинга для общего предка.
Есть варианты формулы со степенью (n+ ni +1) и (n+ ni –1), это в зависимости от того, как считать количество рядов в родословной – до самого пробанда (-1), или до его родителей (+1), математически это равноценные варианты.
Не скажу, что формула плоха. Но! (и это очень большое но) она совершенно не годится в той области, где ее сейчас применяют. Дело в том, что в те времена, никто не знал точно, что же именно является физическим носителем наследственной информации и каким образом происходит передача этой информации от родителей потомству. До открытия ДНК оставалось еще 3 десятилетия. Возможно, и, скорее всего, что Райт считал гены дискретными, не связанными друг с другом частицами, присутствующими в клетке и потому они равномерно перемешиваются при слиянии двух половых клеток. Потому как его формула замечательно описывает процессы смешивания мелких сыпучих веществ и жидкостей.
Давайте рассмотрим на примере. Пусть генами у нас будут кристаллики сахара. Соматическая клетка с полным набором генов будет соответствовать 100 граммам сахара, а половая клетка с половинным набором – 50г. Далее отдельные порции сахара окрашиваем в разные цвета. Смешивание = спаривание.
1 этап. Берем 50г синего сахара (отцовская половая клетка) и тщательно перемешиваем с 50г красного сахара (материнская половая клетка). В результате получаем «щенка» с полным «генотипом» – 100г смеси синего и красного. Издалека выглядит как фиолетовый, вблизи видны синие и красные компоненты, т.е. щенок имеет и признаки своих родителей и в то же время отличается от них. Это щенок имеет 50% генов от папы и 50% от мамы.
2 этап. Наш щенок вырос и тоже стал продуцировать половые клетки, содержащие половинный набор генов, т.е. делим полную порцию пополам, отсыпав 50г сине-красной смеси. Теперь соединяем эту половую клетку с половой клеткой другой собаки, допустим зеленого цвета. В результате мы получаем 100г смеси, где 50% зеленого, 25% синего и 25% красного песку. Синий и красный для этого щенка дед и бабка.
3 этап. Опять отсыпаем 50г сине-красно-зеленой смеси и теперь смешиваем с 50г желтого сахара. Получаем смесь, где 50% желтого, 25% зеленого, 12,5% синего и 12, 5% красного. Синий и красный – прадед, прабабка.
На протяжении 3 этапов мы проводили не родственное скрещивание, но теперь нам захотелось сделать инбридинг и повязать 3-ью с ее синим прадедом.
4 этап. 50г этой пестрой смеси смешиваем с синим сахаром. Получаем: 56,25% синего, 25%желтого, 12,5 зеленого, 6,25% красного. Синий – одновременно и отец, и прапрадед; красный - прапрабабка.
Мы знаем, что особь получает по 50% генов от отца и матери, но в этом случае отец является еще и прапрадедом и потому считается, что у него к отцовским 50% еще прибавляется остаток в 6,25%, который остался в материнской смеси. Вот эти 6,25% и являются коэффициентом инбридинга – это то количество кристалликов сахара, которые совпадают в этой смеси не только по цвету, но и по форме кристалликов. Применительно к собакам мы говорим, что это количество генов, находящихся в гомозиготном состоянии.
А теперь пересчитаем КИ для этого случая по формуле Райта.
Считаем ряды от синего до родителей пробанда:
3 + 0 + 1 = 4;
(1/2)4 = 1/16 = 0,0625;
синий не инбредный, потому fa=0
0,0625 х (1+0)100 = 6,25%.
Все совпадает. Еще бы не совпасть, ведь мы допустили грубейшую ошибку в самом начале – предположили, что гены не связаны друг с другом и комбинируются независимо.
Но, как известно, гены последовательно соединены в линейные цепочки - хромосомы. А, кроме того, все высшие организмы, размножающиеся половым путем, имеют двойной набор хромосом. Давайте возьмем упрощенный пример, когда животное имеет лишь 3 пары хромосом. Предположим что в первой хромосоме сосредоточено 70% генов, во второй – 20%, и в третьей – 10%.
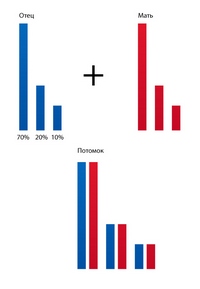
рис. 19
На рис. 19 изображена схема слияния отцовской и материнской половых клеток, в результате чего получился вот такой сине-красный потомок. Это соответствует этапу №1 в опытах с сахаром и, как видим, – соответствует полностью.
Теперь перейдем к этапу №2. С сахаром было все просто – отсыпали 50г и всего делов то. А вот в этом случае, оказывается, есть несколько вариантов получения половых клеток. Имеющийся набор хромосом можно разделить напополам 4-мя способами, получив при этом 8 оригинальных вариантов (рис. 20).
рис. 20 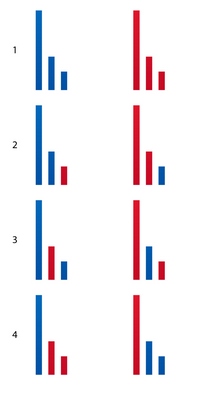
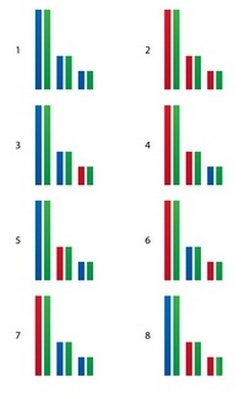
рис. 21
При вязке этих 8-ми особей с зеленым мы можем получить потомков со следующим генотипом (рис. 21).
Согласно опытам с песком и формуле Райта, на этом этапе каждому потомку достается по 25% генов от его деда и бабки. Давайте посмотрим, сколько же их досталось на самом деле:
1й - 50% от деда, 0% от бабки.
2й - 0% от деда, 50% от бабки.
3й - 45% от деда, 5% от бабки.
4й - 5% от деда, 45% от бабки.
5й - 40% от деда, 10% от бабки.
6й – 10% от деда, 40% от бабки.
7й - 15% от деда, 35% от бабки.
8й - 35% от деда, 15% от бабки.
Не существует даже теоретической возможности разделить поровну гены деда и бабки между половыми клетками, чтобы получить искомые 25% у внуков. Возможно, вы скажете, что я специально так хитро задала длину хромосом, чтобы не получалось. Да, специально! Но только лишь по той причине, что такая разница между хромосомами существует на самом деле. Конечно, у собаки не 3, а 39 пар хромосом и генов около 20000 и возможно эти гены распределены по хромосомам так, что существует возможность абсолютно или хотя бы приблизительно точного разделения. Но ведь и количество разных вариантов распределения хромосом при 39 парах несравнимо больше - 239, чем при 3 парах, и равное деление это всего лишь один или несколько вероятных вариантов из этого множества. Так что на этом этапе мы можем лишь сказать, что процент генов деда и бабки в геноме внуков колеблется в пределах от 0 до 50%, но точную цифру мы никак не можем узнать или вычислить. И потому переходить к следующим этапам нет никакого смысла.
Но это еще далеко не все. Давайте посмотрим на особь 1 и 2 с рис. 3, ведь фактически это получились не внуки, а дети одного из предков: синего или красного, при этом каждый из них совсем не родственен другому предку. А ведь рекомбинация и дальше будет происходить подобным образом, и у сине-зеленого потомка, когда начнут вырабатываться половые клетки, то 1/8 их часть будет содержать 3 синие хромосомы, а у красно-зеленого будет такое же количество чисто красных гамет. Получается, что и правнуки синего и красного могут также получить 50% генов своих уже достаточно далеких предков, а это уже получается форменная чушь. Чтобы таких чудес не случалось в реальности, природа предусмотрела дополнительную меру для более активного перемешивания генов предков – кроссинговер. Я коварно ввела всех в заблуждение, когда на рис. 2 изобразила полностью синие и красные хромосомы. На самом деле происходит разрыв цепочек ДНК и обмен отдельными участками (см. рис. 18).
Разрыв может произойти в любом месте ДНК, также возможны множественные перекресты и обмены, а это означает, что количество вариантов перераспределения генов между двумя гомологичными хромосомами, с учетом большой длины хромосом, стремится к бесконечности. После кроссинговера вообще теряется смысл выражений «отцовская хромосома», «материнская хромосома» - теперь каждая хромосома имеет и синие и красные отрезки.
Тот, кто знает математику, сейчас скажет, что элемент хаотичности, который вносит кроссинговер, только на руку нам. Вышесказанная фраза «на этом этапе мы можем лишь сказать, что процент генов деда и бабки в геноме внуков колеблется в пределах от 0 до 50%, но точную цифру мы никак не можем узнать или вычислить», хоть и остается справедливой, но теперь каждый вариант распределения из этого промежутка не равновероятны. Процесс наследования генов от деда или бабки будет описываться кривой нормального распределения или кривой Гаусса (рис. 22).
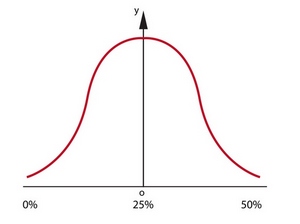
рис. 22
Согласно графику, вероятность того, что потомок получит 0 или 50% генов деда/бабки стремится к нулю, а наибольшую вероятность имеет среднее значение для этого отрезка - 25%.
И что же это получается, что вычисленный по формуле Райта, коэффициент инбридинга хоть и не точно, но с очень высокой степенью вероятности верен? А и опять нет!
Потому, что опять не учтены все физические параметры, описываемой системы. Мы изначально предположили, что хромосома – это не просто цепочка последовательно соединенных генов, но что она состоит только из генов. Однако это не так. В ДНК, состоящих из миллионов нуклеотидов, гены встречаются с той же частотой, что и жизнь в пустыне Сахара. Геном собаки пока не расшифрован, но если учесть, что в геноме человека лишь 1,5% нуклеотидов являются кодирующими последовательностями (генами), а все остальное – мусорная ДНК, и что человеческий геном самый замусоренный в сравнении с другими биологическими видами, то можно приблизительно прикинуть, что у собак гены занимают 2, максимум 3% от общей длины ДНК. Кроме того, гены распределены между хромосомами очень неравномерно. Есть богатые генами хромосомы, а есть практически полностью мусорные.
Но и это еще не все – в геноме собаки есть множество генов абсолютно одинаковых для всех представителей данного биологического вида. Плюс есть некоторое количество генов, которые одинаковы в рамках одной породы (ген бесшерстности у голых собак, ген коротконогости у такс и т.д.).
Так что еще в самом начале, играя с сахаром, мы допустили принципиальную ошибку, окрашивая всю порцию сахара в один цвет. А нужно было в половинной порции 50г брать приблизительно грамм 45 белого + 5г синего и смешивать с красно-белой смесью (45г белого + 5г красного), белые кристаллы сахара – это одинаковые гены в геноме обеих собак, а вот синие и красные – то, что их отличает. А хромосомы следовало бы изобразить в виде длинных белых полосок, на которых изредка кое-где вкраплены синие или красные точки.
При таком малом количестве и такой разбросанности генов на необъятных просторах хромосом, рекомбинация их при производстве половых клеток принимает совершенно непредсказуемый характер. Вычислить, как разойдутся гены нельзя ни точно, ни с какой-то долей вероятности.
Ну и, наконец, последний фактор, вносящий свою долю погрешности. В формуле Райта не учтено, что все гены представлены в геноме в виде аллельной пары и если предок, на которого делают инбридинг, был гетерозиготен по определенному гену, то инбридинг может и не привести к увеличению гомозиготности.
Пример. Пусть наш синенький на самом деле будет черненьким, в смысле окраса и будет нести ген коричневого пигмента – Bb. При вязке с гомозиготным ВВ красным выстрелил сперматозоид с геном b, сине-красный потомок получился Bb и коричневый ген у него точно от отца. Потом мы этого сине-красного вязали с зеленым, который тоже оказался черным гомозиготным и, о чудо (хотя на самом деле никакое не чудо) опять сработала половая клетка с коричневым геном. То же самое повторилось и при вязке с гомозиготным ВВ желтым. Итак, мы получили правнучку синего с генотипом Bb и ген b без вариантов ей достался от прадеда. Потом мы сделали инбридинг, повязав пестренькую с синим, и в этом случае от пестрой участвовала яйцеклетка с геном b, а вот от синего в этот раз выступил сперматозоид с геном B.
В опытах с сахаром и в рисунках хромосом мы бы посчитали эту пару генов одинаково синими, т.е. гомозиготными, а на самом деле это самая что ни на есть гетерозиготная пара.
Вывод: заниматься вычислениями КИ это бесполезное и бессмысленное занятие. Та информация, которую нам дает этот коэффициент, можно высказать простыми человеческими словами: инбридинг 1:2, 2:1, 2:2 это очень близкий инбридинг и не стоит его применять без архиважной надобности, а также и не следует в пределах одной родословной накапливать слишком много инбридингов на одних и тех же собак.
Если тесный инбридинг такая неприятная вещь, то не стоит ли вообще от него отказаться и даже запретить использовать в разведении? В этом есть рациональное зерно, однако не стоит выплескивать вместе с водой и младенца. Даже от кровосмешения может быть польза. Кобеля, которого известные специалисты оценивают как восходящую звезду, прочат большое будущее, а владельцы невест заранее резервируют очередь на вязку, желательно бы проверить в близком инбридинге. Далеко не все наследственные заболевания сейчас тестируются в лабораториях и если удастся селекционным методом как можно раньше выявить наличие вредной мутации в геноме кобеля, то это позволит избежать того колоссального вреда, который можно было нанести породе, используя такого неблагополучного производителя. Второй плюс – щенки, полученные в тесном инбридинге на выдающегося представителя породы, являются своеобразным хранилищем генетических последовательностей своего предка. Да, сами они могут быть хуже своего знаменитого родителя, но зато могут оказаться весьма ценными производителями.
| <== предыдущая лекция | | | следующая лекция ==> |
| Требования к качеству сладких блюд | | | Что такое атеросклероз |
Дата добавления: 2018-11-26; просмотров: 1113;
Поиск по сайту
Узнать еще
- III. ТРЕБОВАНИЯ РКФ ДЛЯ ДОПУСКА СОБАК В ПЛЕМЕННОЕ РАЗВЕДЕНИЕ
- Вопрос 77. Генокопии и фенокопии в патологии человека. Критика представлений о фатальности наследственных заболеваний. Евгенетика
- Генетика и эволюция. Молекулярные механизмы наследственности и изменчивости
- Групповая замкнутость и социальная селекция
- История собаководства в дореволюционной и современной России. Структура собаководства в России. История прикладного собаководства и собакозаводчества на Руси
- Крупномасштабная селекция в скотоводстве
- Лекция № 11-12: Генетика пола. Скрепленное с полом наследование. Влияние внешней среды на генотип. Предупреждение наследственных заболеваний человека.
- Лекция №14: Селекция растений и животных. Мутагенез. Достижения в селекции животных и растений в РК.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
