КОВАЛЕНТНАЯ МОДИФИКАЦИЯ ФЕРМЕНТОВ И ЕЕ ТИПЫ
Ковалентная модификация ферментов – тип регуляции ферментативной активности под действием других ферментов, осуществляющийся за счет присоединения к белковой молекуле небольших химических групп, что вызывает конформационные изменения в молекуле белка, приводящие к изменению определенной геометрии каталитического центра. Является быстрым механизмом регуляции активности ферментов внешними сигналами. Ковалентная модификация является основным механизмом при рецептор-зависимом ответе клеток на внешние воздействия, тогда как аллостерические эффекторы изменяют активность ферментов в ответ на изменение внутриклеточных условий.
Фосфорилирование – наиболее распространенный тип ковалентной модификации, является АТФ-зависимым процессом и осуществляется с помощью ферментов протеинкиназ. В большинстве случаев фосфорилированная форма фермента более активна, чем нефосфорилированная. Фосфорилирование модифицирует белки добавлением отрицательно заряженных остатков фосфорной кислоты к гидроксильным группам остатков серина, треонина и, реже, тирозина.
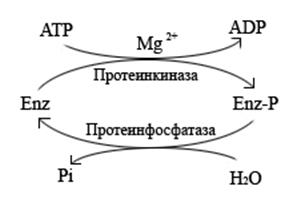
Рис. 4.3.1. Фосфорилирование/дефосфорилирование.
Возвращение фермента в исходное состояние осуществляется ферментами протеинфосфатазами, которые отщепляют неорганический фосфат от молекулы белка.
В некоторых случаях фосфорилирующие и дефосфорилирующие белки сами являются модифицируемыми ферментами. Активность протеинкиназ и протеинфосфатаз находится под гормональным контролем и регулируется нервной системой.
К ковалентной модификации ферментов помимо фосфорилирования относятся также аденилирование, уридилирование, ацетилирование, АДФ-рибозилирование.
Ковалентную модификацию аденилированием и уридилированием можно рассмотреть на примере регуляции активности фермента глутаминсинтетазы. Глутаминсинтетаза выполняет одну из ключевых функций в обмене азота у бактерий E. Coli. При выращивании Е. coli на среде, в которой источником азота служит не NH4+, а другие соединения, в клетках наблюдается высокий уровень глутаминсинтетазной активности. Добавление в ростовую среду солей NH4+ вызывает быстрое (в течение минуты) десятикратное снижение активности фермента, которое нельзя объяснить подавлением его синтеза. Попытки объяснить данное явление привели к открытию фермента, катализирующего инактивацию глутаминсинтетазы in vitro в присутствии АТФ, Mg2+ и глутамина. Далее было установлено, что разные формы фермента содержат различные количества ковалентно связанной АМФ. Далее было установлено, что инактивирующий белок является аденилилтрансферазой, катализирующей ковалентное присоединение АМФ к определенной тирозильной группе в каждой из 12 субъединиц глутаминсинтетазы. Однако полное превращение происходит только в экстремальных условиях, например когда клетки, выращенные в условиях недостатка NH4+, попадают в среду, богатую этим соединением. Обычно глутаминсинтетаза находится в промежуточных состояниях аденилирования, в которых аденилированными являются лишь некоторые из 12 субъединиц.
Активность отдельной субъединицы в составе олигомера зависит от состояния соседних субъединиц (аденилированы они или не аденилированы), вероятно, из-за аллостерического влияния, которое передается через межсубъединичные контакты.
При переносе Е. coli из среды, богатой NH4+, в бедную этим соединением среду происходит быстрая реактивация глутаминсинтетазы, что указывает на обратимость процесса аденилирования. Было установлено, что активность аденилилтрансферазой регулируется с помощью уридилирования и деуридилирования. Уридилирование происходит в результате катализируемого специфичной уридилилтрансферазой ковалентного присоединения УМФ к одному или двум тирозильмым остаткам каждой субъединицы аденилилтрансферазы. Деуридилирование катализируется уридилил-отщепляющим ферментом. Уридилирующий и деуридилирующий ферменты образованы одним полипептидом, который, подобно аденилилтрансферазе, проявляет бифункциональные свойства.
Примером ковалентной модификации белков с помощью ацетилирования является модификация гистонов. Ацетилирование гистонов играет важную роль в модуляции структуры хроматина при активации транскрипции, увеличивая доступность хроматина для транскрипционного аппарата. Гистоны целенаправленно модифицируются на тех промоторах, которые требуется активировать. Ацетилированию и деацетилированию подвергаются определенные остатки лизина, что осуществляется с помощью ферментов ацетилтрансфераз и деацетилаз. Предполагается, что ацетилированные гистоны менее прочно связаны с ДНК и поэтому ацетилирование может облегчать доступ и связывание факторов транскрипции с их элементами узнавания на ДНК. Ацетилирование остатков лизина в N-концевых «хвостиках» гистонов H2A, H2B, H3 и H4 нейтрализует их положительный заряд и соответственно блокирует ассоциацию с витками нуклеосомной ДНК. Это, в свою очередь, декомпактизует структуру как самой нуклеосомы, так и хроматина в целом и освобождает внешнюю поверхность витков ДНК для взаимодействий с регуляторными факторами.
Еще одним способом химической модификации ферментов с помощью присоединения небольших химических групп является АДФ-рибозилирование. АДФ-рибоза может вызывать неэнзиматическое АДФ-рибозилирование ряда белков. К ним относятсябелки регулирующие репарацию ДНК, белки-регуляторы трансляции и агрегации тромбоцитов. Моно-АДФ-рибозилирование – посттрансляционный процесс модификации белков, ответственный за токсический эффект некоторых бактериальных токсинов. Действие коклюшного или холерного токсинов вызывает моно-АДФ-рибозилирование белков плазматической мембраны и может изменять работу сигнальных систем или активность мембранного транспорта в результате нарушения функционирования G-белков. Схематически действие холерного токсина можно описать следующим образом (Рис. 4.3.2). В норме ГТФ, связанный с Gsα быстро гидролизуется (синяя стрелочка), так, что активация аденилатциклазы и увеличение цАМФ происходит столь долго, сколько гормон связан с рецептором. В присутствии холерного токсина Gsα необратимо модифицируется АДФ- рибозилированием, так, что она может связывать ГТФ, но не может его гидролизовать (красные стрелочки). В результате постоянной активации происходит нерегулируемый рост уровня цАМФ.
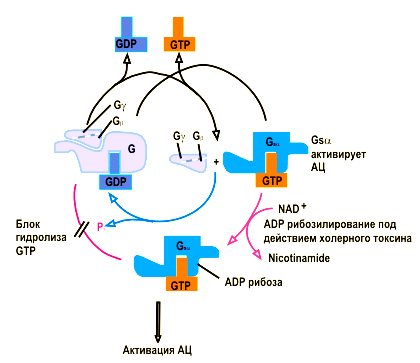
Рис. 4.3.2. Механизм действия холерного токсина
При нормальных условиях АДФ-рибозилированию могут подвергаться актин, десмин, интегрин-α-7.
Рис. 4.3.3. АДФ-рибоза
Аденилатциклаза
Аденилатциклаза (АТФ -пирофосфатлиаза циклизирующая) фермент, класса лиаз, катализирует гидролиз АТФ с образованием цАМФ и пирофосфата.
Mg2+ АТФ à цАМФ + H4P2O7
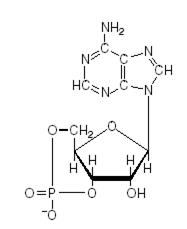
Рис. 4.3.4. структура цАМФ
Аденилатциклаза обнаружена практически во всех тканях животных, а также у бактерий. В клетках животных он локализован в плазматических мембранах, у бактерий – в мембранах и цитоплазме. Наиболее существенная роль цАМФ состоит в активации цАМФ – зависимых протеинкиназ (протеинкиназы А).
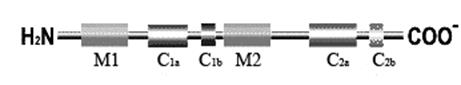
Рис. 4.3.5. Первичная структура аденилатциклазы
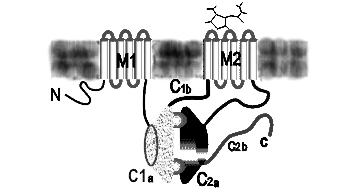
Рис. 4.3.6. Расположение аденилатциклазы в мембране
Фосфолипаза С
Фосфолипаза С является ключевым ферментом фосфоинозитидной мессенджерной системы. Многочисленные внеклеточные сигнальные молекулы (гормоны, нейромедиаторы, факторы роста, иммуноглобулины, антигены и др.), при взаимодействии со своими рецепторами вызывают активацию фосфолипазы С (ФЛC). При взаимодействии лиганда с рецептором активурующий ФЛC сигнал может передаваться специальным G-белком (Gq). Активированная ФЛC катализирует расщепление мембранного фосфолипида фосфатидилинозитол-4,5-дифосфата (ФИФ2) на инозитолтрифосфат (ИФ3) и диацилглицерол (ДАГ). Диацилглицерол связывается и стимулирует протеинкиназу С (ПKC), ИФ3 взаимодействует с активируемым им Са2+ каналом (ИФ3 рецептором), в результате чего происходит выход ионов кальция из эндоплазматического ретикулума, которые участвуют в активации Са2+ - кальмодулин-зависимых протеинкиназ, протеинкиназы С и многочисленных кальций-чувствительных белков. Таким образом, в отличие от других ферментов, катализирующих образование вторичных мессенджеров, ФЛС использует компоненты самой мембраны.
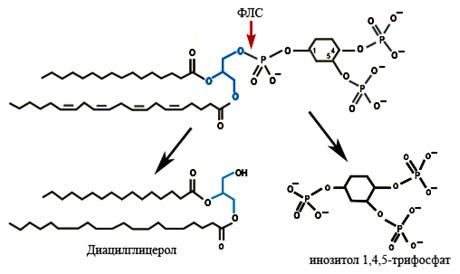
Рис. 4.3.7. Действие фосфолипазы С на фосфатидилинозитол-4,5-дифосфат
Дата добавления: 2020-10-01; просмотров: 1105;
Поиск по сайту
Узнать еще
- ОСНОВНЫЕ ТИПЫ И СВОЙСТВА НАПОЛЬНЫХ И БОРТОВЫХ СИСТЕМ ТЕХНИЧЕСКОГО ДИАГНОСТИРОВАНИЯ
- II.II. Типы геологических карт
- IV.I. Ловушки нефти и газа и их типы
- Pис. 60. Типы складок по форме замка: а - обычная открытая. б - остроугольная, в - тупая складка
- А.1. Назначение и типы сборных элементов и конструкций
- Авто- и гетеростереотипы русских
- Адресная арифметика, типы указателей и операции над ними
- Аллергия. Типы аллергических реакций и принципы их терапии

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
