Температура, освещенность и растворенный кислород как ключевые экологические факторы водотоков
A. Температура. Температура представляет собой одну из наиболее значимых переменных, определяющих экологические процессы в биосфере. Данный физический параметр напрямую воздействует на движение молекул, динамику жидкости, константы насыщения газов, растворенных в воде, и скорость метаболизма организмов. Многочисленные дополнительные факторы, влияющие на жизнь на Земле, также зависят от теплового режима, что делает его основополагающим для функционирования экосистем. В проточных водах солнечное излучение обычно служит главным источником тепла, особенно там, где прямые солнечные лучи покрывают большую часть водной поверхности.
Регуляция температуры воды происходит под действием множества факторов в различных пространственных масштабах. На малых лесных ручьях густой растительный полог затеняет водную поверхность, эффективно поддерживая прохладный тепловой режим. Однако вырубка прибрежных лесов и раскрытие полога могут значительно повысить температуру воды в ручьях (Джонсон и Джонс, 2000). Кроме того, теплопередача от грунтовых вод играет исключительно важную роль в регулировании тепловых режимов (Бакстер и Хауэр, 2000; Меллина и др., 2002). Независимо от конкретных механизмов контроля, температура оказывает глубокое влияние на присутствие, отсутствие, жизненный цикл и пространственное распределение водных организмов (Хокинс и др., 1997; Лоу и Хауэр, 1999; Хауэр и др., 2000; Эберсоле и др., 2001).
Годовые колебания температуры воды в водотоке имеют критическое значение для водной биоты. Ключевые параметры жизненного цикла, включая размножение и рост водных растений и животных (от диатомовых водорослей и водных насекомых до рыб и других пойкилотермных позвоночных), регулируются тепловыми условиями. Многие обитатели ручьев используют температуру или её изменения в качестве экологического сигнала для выхода из куколок (у насекомых) или нереста (у рыб). В регионах с умеренным климатом зимние температуры воды могут приближаться к 0°C, тогда как летом превышать 30°C (Хауэр и Бенке, 1986; Лоу и Хауэр, 1999).
В ручьях обычно наблюдаются суточные перепады температур, при этом дневные диапазоны часто превышают 5°C (рис. 5.1).
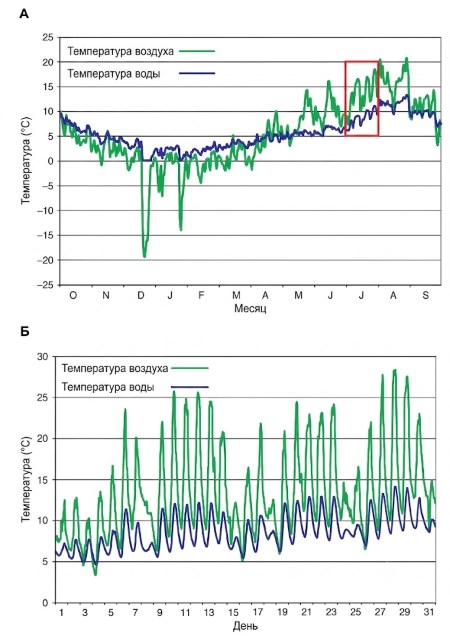
Рисунок 5.1. (А) Среднесуточные температуры воздуха и воды, рассчитанные по среднечасовым значениям (°C), Макдональд-Крик, национальный парк Глейшер, Монтана (США), с 1 октября 1998 г. по 30 сентября 1999 г. (B) Среднечасовые температуры воздуха и воды, рассчитанные с интервалом в 5 минут (мгновенные измерения, °C), Макдональд-Крик, национальный парк Глейшер, Монтана (США), июль 1999 г. (красная рамка на панели A).
В особых условиях возможны исключительно высокие колебания температуры. Например, очень маленькие альпийские ручьи, получающие прямое солнечное излучение, в конце лета могут днём прогреваться выше 20°C, а ночью остывать почти до 0°C. Даже на крупных реках с расходом воды более 500 м³/с наблюдаются суточные перепады в диапазоне 3–5°C. Тем не менее, благодаря высокой скрытой теплоте воды (для изменения температуры даже на 1°C требуется значительное поглощение или выделение энергии), ежедневные колебания температуры воды в потоке обычно гораздо уже, чем у воздуха.
Зимой ручьи и реки в умеренных, арктических и горных районах часто замерзают, что существенно влияет на расход воды, проникновение света, концентрацию растворённого кислорода и множество других факторов. Образование льда может серьёзно нарушить среду обитания и повлиять на распределение и поведение водных организмов (Брэдфорд и др., 2001). Такая зимняя динамика представляет собой важное сезонное препятствие для водной флоры и фауны холодных регионов.
Данные о температуре в пределах одного водотока часто приводятся в виде режима для одной точки, что порождает ошибочное мнение об однородности теплового режима во всех местообитаниях. Напротив, температура воды может сильно различаться на участках, разделённых всего несколькими метрами. Зоны подруслового потока (гипорейная зона) часто намного теплее, чем воды в основном русле, особенно в аллювиальных реках с гравийным дном, где велика связь между русловыми и грунтовыми водами. Местообитания, питаемые грунтовыми водами (см. главы 6 и 33), летом могут быть на несколько градусов холоднее, а зимой – теплее основного русла. В водотоках также наблюдаются значительные продольные перепады температур – от затенённых истоков до широких открытых участков. Это явление особенно выражено в горных районах, где река берёт начало в альпийских условиях, а затем протекает в более тёплом климате ниже по течению, прежде чем впасть в другие воды, озеро или океан (Хауэр и др., 2000).
B. Освещённость. Освещённость является критической переменной в большинстве экосистем, а в водотоках солнечная радиация необходима для фотосинтеза водорослей и макрофитов. Солнечный свет также обеспечивает возможность визуального поведения (например, хищничество рыб). Поскольку ручьи тесно связаны с окружающими наземными ландшафтами, на режимы освещённости сильно влияют прибрежные объекты, такие как деревья или геологические структуры. Тень от нависающих крон ограничивает первичную продуктивность многих нетронутых лесных ручьёв (например, Хилл и др., 1995). Продольное (вниз по течению) изменение светового режима и его последствия для биоэнергетики ручья являются неотъемлемой частью динамики экосистемы (Ваннот и др., 1980). Даже в регионах, где прибрежная растительность даёт мало тени, крутые берега или стены каньона могут значительно уменьшить количество света, достигающего русла (Миншолл, 1978).
Хотя длины волн солнечного излучения варьируются от <300 до >5000 нм, для водных экологов наибольший интерес представляет диапазон 400–700 нм, связанный с процессами фотосинтеза. Этот спектральный интервал называется фотосинтетически активным излучением (ФАР, PAR). Он примерно соответствует диапазону, воспринимаемому человеческим глазом (видимому свету). ФАР измеряется с помощью квантовых датчиков – фотоэлементов, определяющих количество фотонов в диапазоне 400–700 нм, попадающих на единицу площади за единицу времени. Эта величина называется плотностью потока фотонов (PFD). Единицы измерения PFD (и PAR) – мкмоль квантов·м⁻²·с⁻¹ или мкЭйнштейны·м⁻²·с⁻¹. Эйнштейн (моль фотонов) не является единицей СИ, и его использование сокращается. В экологических исследованиях, охватывающих длительные периоды, PAR часто выражают как моль квантов·м⁻²·сут⁻¹.
Плотность потока фотонов в речных экосистемах сильно меняется во времени. В солнечный день PFD в незатенённых водотоках может колебаться от 0 (перед рассветом или после заката) до >2000 мкмоль квантов·м⁻²·с⁻¹ (в полдень). Облачность увеличивает внутридневную изменчивость, а прохождение погодных фронтов создаёт значительные суточные вариации. Сезонные колебания светового режима обусловлены изменением угла наклона Солнца, продолжительности дня и фенологическими изменениями прибрежной растительности. Например, эффект затенения от весеннего распускания листьев в лиственных лесах снижает уровень PFD на поверхности ручьёв на два порядка (Хилл и др., 2001).
Пространственная изменчивость освещённости также высока. Количество тени, отбрасываемой растительностью на берегах, в значительной мере определяет пространственные различия. Разрывы в кронах деревьев над лесными ручьями создают существенную мозаичность освещённости от участка к участку. Одним из наиболее важных антропогенных воздействий на водные экосистемы является изменение светового режима вследствие удаления прибрежной растительности. Прибрежная растительность также играет ключевую роль в продольном градиенте интенсивности освещения в ненарушенных системах. По мере увеличения размера ручья ниже по течению прибрежные деревья и кустарники затеняют пропорционально меньшую часть потока, позволяя большему количеству прямых и рассеянных солнечных лучей достигать русла.
Когда свет достигает поверхности воды, он ослабляется самой водой и, что ещё важнее, веществами, растворёнными или взвешенными в ней. В чистых и неглубоких водотоках это ослабление незначительно, поэтому интенсивность света на поверхности достаточно точно отражает таковую у дна. Однако в некоторых водотоках наблюдается значительное количество взвеси или они окрашены растворёнными органическими веществами (РОВ). Проникновение света в такие потоки сильно снижено, особенно на глубоких участках, поэтому поверхностные измерения не отражают условий для биоты в русле. Подводные измерители освещённости необходимы для определения светового режима в мутных или сильно окрашенных потоках. Ослабление света в них описывается экспоненциальной функцией, аналогичной используемой лимнологами и океанографами: E(z) = E(0) · e⁻ᵏᶻ, где E(z) и E(0) – освещённость на глубине z и на поверхности соответственно, а k – коэффициент затухания (Кирк, 1994).
За последние 15 лет всё большее внимание уделяется роли ультрафиолетового излучения (длина волны <400 нм) в водных экосистемах. Как УФ-А (320–400 нм), так и УФ-В (280–320 нм) оказывают вредное воздействие на речные водоросли и беспозвоночных (например, Ботвелл и др., 1994; Келли и др., 2003) в чистых, незатенённых, неглубоких водотоках. Ультрафиолетовый свет в значительной степени поглощается растворённым органическим веществом, что вызывает интерес как к защитной функции РОВ, так и к эффекту УФ-излучения, делающему РОВ более доступным для бактериального метаболизма (например, Бриско и Циглер, 2004).
C. Кислород. Растворённый кислород (DO) непосредственно влияет на водную флору и фауну через его доступность и метаболизм, а также косвенно воздействует на различные биогеохимические процессы. В большинстве незагрязнённых водотоков и рек концентрация DO остаётся значительно выше 80% насыщения. Растворимость кислорода нелинейно возрастает при понижении температуры и уменьшается с падением атмосферного давления (связанным с высотой над уровнем моря или барическими изменениями погоды). Эти физико-химические взаимосвязи имеют большое значение для распределения водных организмов и физиологии дыхания.
Концентрации растворённого кислорода неодинаковы в пределах водотока и между его истоками. Поступление подземных вод из гипорейной зоны (см. главы 6 и 33) или боковой поток грунтовых вод могут создавать участки дна с концентрацией кислорода значительно ниже, чем в окружающей воде. Почти все речные организмы чувствительны к уровню DO. Органические загрязнения, например, связанные со сбросом городских сточных вод или промышленных отходов, способны существенно снизить концентрацию DO во всём водотоке, поскольку микробиологические процессы поглощают кислород из воды. Это явление называют биохимической потребностью в кислороде (БПК).
В незагрязнённых проточных водах концентрация кислорода также может сильно различаться в зависимости от микроместообитания. Активность микроорганизмов в скоплениях листьев и детрита способна снижать локальный уровень DO. В ручьях и реках с обильным развитием водорослей наблюдаются большие суточные колебания кислорода: фотосинтез повышает его концентрацию днём, а дыхание снижает ночью. Для оценки валовой первичной продуктивности, дыхания и чистой первичной продуктивности в водотоках используют исследования метаболизма всей экосистемы на основе изменений содержания кислорода (Малхолланд и др., 2001). Эти переменные, а также экологическое значение отношения продукции к дыханию (P:R), более подробно обсуждаются далее.
Сведения об авторах и источниках:
Авторы: Ф. Ричард Хауэр и Гэри А. Ламберти
Источник: Методы в экологии ручьев
Данные публикации будут полезны студентам экологических и географических специальностей, начинающим специалистам в области гидрологии, геоморфологии и управления водными ресурсами, а также всем, кто интересуется динамикой речных экосистем и вопросами охраны природных ландшафтов.
Дата добавления: 2026-05-20; просмотров: 243;
Поиск по сайту
Узнать еще
- ChatGPT в образовании: как интегрировать искусственный интеллект в учебный процесс, развивать цифровую грамотность и преподавать этику ИИ для будущих специалистов
- ChatGPT для преподавателей: как использовать функции ИИ в обучении (практическое применение, преимущества, ограничения)
- ChatGPT для учителей: как автоматизировать административные задачи, отчетность и коммуникацию для повышения эффективности работы
- ChatGPT для учителей: как использовать ИИ для непрерывного профессионального развития (НПР) и карьерного роста
- Административная ответственность за экологические правонарушения: основания, меры и порядок применения
- Аномалия плотности воды: почему лед не тонет и как это явление спасает жизнь на Земле
- Атмосферный фронт как поверхность раздела воздушных масс
- Биология как наука: предмет изучения, методы и основные направления

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
