Типы замен оснований в молекулах ДНК
| Исходное основание | Основание, занявшее место исходного | Тип замены |
| Пурин | Другой пурин | Простая замена (транзиция) |
| Пиримидин | Другой пиримидин | То же |
| Пурин | Любой пиримидин | Перекрестная замена (трансверсия) |
| Пиримидин | Любой пурин | Перекрестная замена (трансверсия) |
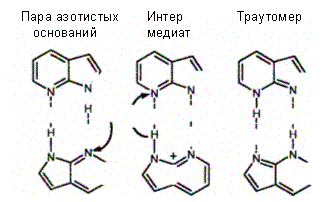
Спонтанные замены азотистых оснований происходят очень редко. Например, в соответствии с существующими расчетами у человека за год случается около 10—20 спонтанных замен оснований, причем одна замена может быть повторена на каждые 10 000 генов лишь 50 раз на протяжении времени в 1 млн лет. Можно полагать, что такая чрезвычайно низкая частота замен оснований в ДНК присуща как животным (млекопитающим), так и растениям. Спонтанные замены азотистых оснований возникают в ДНК в результате «ошибок», совершаемых ДНК-полимеразой и сопровождающихся неправильным спариванием оснований. Одно из объяснений этой «ошибочности» было дано Д. Уотсоном и Ф. Криком еще в 1953 г. и оно сводится к признанию в ошибочном спаривании роли тауто-мерных форм (структур, в которых протон перешел на место, противоположное обычной водородной связи) естественных оснований. Следовательно, структурные основы для мутаций в виде замен оснований обеспечивают таутомеры естественных оснований.
 Транзиции индуцируются азотистой кислотой, которая вызывает окислительное дезаминирование аденина, цитозина и гуанина, содержащих свободные аминогруппы, в гипоксантин, урацил и ксантин соответственно. Из-за того, что дезаминирование сопровождается переходом аминооснования в кетонооснование, гипоксантин, например, подобно гуанину, будет спариваться с цитози-ном, т. е. в результате дезаминирования аденина в гипоксантин пара А-Т перейдет в пару Г-Ц. В случае дезаминирования цитозина в урацил пара Г-Ц перейдет в пару А-Т. Транзиции индуцируются также алкилирующими соединениями. Например, этилметан-сульфонат алкилирует гуанин и освобождает от него ДНК без нарушения ее сахарофосфатного каркаса. Следовательно, гуанин может быть заменен любым основанием, и это ведет не только к транзициям, но и к трансверсиям.
Транзиции индуцируются азотистой кислотой, которая вызывает окислительное дезаминирование аденина, цитозина и гуанина, содержащих свободные аминогруппы, в гипоксантин, урацил и ксантин соответственно. Из-за того, что дезаминирование сопровождается переходом аминооснования в кетонооснование, гипоксантин, например, подобно гуанину, будет спариваться с цитози-ном, т. е. в результате дезаминирования аденина в гипоксантин пара А-Т перейдет в пару Г-Ц. В случае дезаминирования цитозина в урацил пара Г-Ц перейдет в пару А-Т. Транзиции индуцируются также алкилирующими соединениями. Например, этилметан-сульфонат алкилирует гуанин и освобождает от него ДНК без нарушения ее сахарофосфатного каркаса. Следовательно, гуанин может быть заменен любым основанием, и это ведет не только к транзициям, но и к трансверсиям.
Транзиции часто вызываются мутагенами, действующими на ДНК только в состоянии репликации, например, 5-бромурацилом, который является аналогом тимина и способен включаться в ДНК посредством замещения тимина. Наряду с нормальной способностью 5-бромурапила спариваться с аденином иногда возникает состояние, когда он действует не как тимин, а как цитозин, что обеспечивает формирование водородных связей его не с аденином, а с гуанином. Эти «ошибки» спаривания происходят либо при включении 5-бро-мурацила в ДНК («ошибки» включения), либо при репликации ДНК после его включения («ошибки» репликации). Следовательно, время «ошибок» определяет характер транзиции. «Ошибки» спаривания, индуцируемые 5-бромурацилом, ведут к транзициям от пары Г-Ц к паре А-Т, и наоборот (от А-Т к Г-П). Подобные транзиции индуцируются также 2-аминопурином.
Замены оснований приводят к изменениям смысла кодонов, вследствие чего они приобретают способность кодировать другую аминокислоту (миссенс-мутации). Например, замена в триплете ГУА, содержащемся в гене р-гемоглобина, урацила на аденин (трансверсия) сопровождается тем, что в цепи р-гемоглобина вместо валина оказывается глутаминовая кислота. Это ведет к превращению гемоглобина в новый вариант мутантного гемоглобина (например, типа Бристоль). В результате замен оснований возникают также нонсенс-мутации, когда на измененных кодонах обрывается чтение информации гена (как правило, такими кодонами являются триплеты УАГ, УАА и УГА). Одновременно в результате замен образуются кодоны, сохраняющие исходный смысл.
Делеции и включения одного или нескольких азотистых оснований в нуклеотидных последовательностях ДНК могут быть ошибками репликации ДНК или индуцироваться акридиновыми красителями. Такие изменения называют мутациями сдвига рамки, ибо они приводят к сдвигу «рамки чтения» кода гена. Включаясь между соседними основаниями, акридин оранжевый заставляет их «раздвигаться» на расстояние в 0,6-0,8 нм.
Если акридин оранжевый присутствует в полинуклеотидной цепи-шаблоне, то результатом будет добавление основания в новую цепь в процессе репликации ДНК. Если же акридин оранжевый присутствует в клетке во время репликации ДНК, то он может включаться в новую цепь вместо основания, имитируя парное (противоположное) основание в цепи-шаблоне, и затем выйти. Это приводит к тому, что вновь реплицированной цепи будет недоставать основания, т. е. она будет реплицирована с делецией по основанию- Делеции могут затрагивать несколько оснований. Например, описаны делеции 15 оснований, которые сопровождались утратой в белке 5 аминокислот.
Дупликации (добавление) 1—2 оснований могут приводить также к мутациям со сдвигом «рамки считывания» кода. Если дупли-кация происходит внутри гена, то «рамка считывания» нарушается на большом протяжении.
Делеции и дупликации азотистых оснований представляют собой молекулярный механизм и мутации митохондриальной ДНК человека. Установлено, что из мтДНК человека могут быть делегированы сегменты длиной до 5000 пар оснований.
Особую форму молекулярных механизмов генных мутаций представляют повторы триплетов азотистых оснований. Наличие в молекулах ДНК повторов триплетов оснований сопровождается нарушениями нормального цикла репликации ДНК, с одной стороны, и аномальным синтезом белка (из-за повторов аминокислоты, кодируемой повторяющимся триплетом), с другой стороны. Например, мутации гена, контролирующего белок хантингтан, недостаток которого у человека сопровождается болезнью Хантингтона, заключаются в резком увеличении повторов триплета ЦАГ.
Дата добавления: 2016-05-30; просмотров: 2697;
Поиск по сайту
Узнать еще
- I. Политический режим: понятие, сущность и основные типы.
- I. Понятие и типы политических партий.
- II. Типы политических лидеров.
- MS Word: проверка орфографии, поиск и замена, режимы просмотра документа (обычный, разметки страницы, структура).
- VI. ЩЕЛЕВЫЕ И СПЕЦИАЛЬНЫЕ ТИПЫ АНТЕНН
- А – трехгранный; б – в форме ложечки; в – плоский с заменяемой пластиной из твердого сплава
- А) Типы и конструктивные особенности баз.
- А. Клинические типы.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
