МОРСКАЯ МИКРОБИОЛОГИЯ
Интерес человека к обитателям моря традиционен. В концеД прошлого века стало известно, что бактерии развиваются и b| морской воде. Однако оказалось в некотором роде сенсационным открытие, сделанное в 1884 г. французским микробиологом М. Серте, участником биологической экспедиции на судне «Талисман» в Атлантический океан. Он обнаружил, что бактерии,] минерализующие органические вещества, присутствуют нетолько в поверхностных слоях воды, но и на глубине в несколько тысяч метров. Большую роль в развитии морской микробиологии сыграли исследования немецкого микробиолога Б. Фишера, участника экспедиций, организованных знаменитым географом и натуралистом А. Гумбольдтом. В конце 80-х годов прошлого! века Б. Фишер установил, что прибрежные районы морей богаче бактериями, чем воды открытого океана, и что наибольшее число бактерий обитает на глубинах 200—400 м. Он .обнару-' жил также, что для развития многих морских форм бактерий ' необходимо присутствие в среде соли.
Особое значение для развития морской микробиологии в начале нашего века имели работы Бориса Лаврентьевича Исаченко (1871—1948)—одного из создателей морской микробиологии. В 1906 г. он принимал участие в экспедиции в Баренцево море и в Северный Ледовитый океан. Им впервые были полу- • чены данные о развитии бактерий, относящихся к различным физиологичским группам, в морской воде и осадках. Б. Л. Исаченко пришел к выводу о разносторонней геохимической деятельности морских микроорганизмов. Его монография «Микробиологические исследования Северного Ледовитого океана» (1914) получила всемирное признание. В 20-е—40-е годы Б. Л. Исаченко исследует микрофлору Черного, Мраморного, Азовского, Карского морей и моря Лаптевых. При изучении морских осадков в районе впадения в Карское море Енисея и Оби им было показано, что по месту нахождения на дне моря спор В. mycoides можно определить районы проникновения пресных вод в океан, поскольку эта бактерия типична только для микрофлоры почвы и попадает в море с речной водой.
Г. А. Надсоном проведены исследования зеленых и бесцветных серобактерий в Баренцевом, Балтийском, Черном и Каспийском морях. Ряд его работ посвящен изучению физиологии светящихся бактерий.
В 1930—1939 годы в Морском научном институте (ныне Всесоюзный НИИ рыбного хозяйства и океанографии) бактериологической лабораторией заведывал Владимир Степанович Буткевич (1872—1942), принимавший участие в нескольких экспедициях в Баренцево, Каспийское и Азовское моря. Он установил, что на границе Гольфстрима в Северном Ледовитом • океане значительно возрастает численность микроорганизмов.
В С. Буткевнч исследовал роль бактерий в процессах денитри-сЬикации в море, в отложении железо-марганцевых конкреций, в "щевых цепях морских организмов. Им предложен микробиологический метод разведки газов и нефти.
В 1933 г. микробиологическая лаборатория организуется на Севастопольской биологической станции (ныне Институт биологии южных морей АН УССР). Благодаря работам сотрудников этой лаборатории стали известны закономерности распространения микроорганизмов в морях Средиземноморского бассейна Атлантического и Индийского океанов, изучена роль микроорганизмов в продукционных процессах. В 40-е годы начинаются микробиологические исследования на базе Мурманской биологической станции (ныне Мурманской морской биологический институт Карельского филиала АН СССР). Главное внимание зг'есь уделяется микробиологическим процессам в зоне приливно-отливных вод.
В 1950 г. в Институте микробиологии АН СССР в Москве был организован отдел морской микробиологии, который возглавил Анатолий Евсеевич Крисе (1908—1984). Основное на* правление работ этого отдела было посвящено изучению эколого-географического распределения бактерий в океанах. Установлена индикаторная роль бактерий в обнаружении течений, рассчитана биомасса и продукция бактерий в открытых районах океана, исследована микрофлора воды в районе Северного Полюса и в антарктических морях, проведено изучение действия высокого гидростатического давления на бактериальную клетку. Основные результаты этих работ изложены в монографиях А. Е. Крисса «Морская
микробиология (глубоководная)» (1959), «Микробное население Мирового океана (видовой состав, географическое распространение)» (1964), «Жизненные процессы и гидростатическое давление» (1973), «Микробиологическая океанография» (1976).
Большое значение развитию исследований в области морской микробиологии придают во многих странах, имеющих выход к морям.
Во Франции исследования микробиологии моря сосредоточены в Марселе, где изучаются систематика морских микроорганизмов, их взаимоотношения с водорослями и "беспозвоночными, их роль в процессах самоочищения моря и т. д.
- В ФРГ центр по исследованию морских организмов организован в Киле, в Великобритании—в Плимуте и в Шотландии.
В США микробиологические исследования моря были начаты в Скриппсовском институте в Да Холле (Калифорния) и в Вудс Холе (Массачусетс). Особенно большой вклад в развитие морской микробиологии был внесен работами К. Зобелла, в 1946 г. опубликовавшего книгу, ставшую классическим руководством по морской микробиологии.
В Японии при Токийском университете в 1966 г. создан отдел морской микробиологии, который возглавил проф. Н. Тагг Микробиологические лаборатории созданы в Японии во всекрупных университетах и институтах при центрах марикультур во многих префектурах.
В последние годы интерес к морской микробиологии резко возрос в связи с сенсационными находками жизни бактерий: глубоководных горячих источниках на дне океана и открытие явлений симбиоза морских животных с хемоавтотрофными бактериями.
ГЕОМИКРОБИОЛОГИЯ
Геомикробиология является одним из разделов экологиче ской микробиологии. В ее задачи входит выяснение роли микроорганизмов в процессах миграции, рассеяния и накопления .химических элементов в биосфере. Геомикробиология тесня смыкается с другими разделами экологической микробиологии—водной и почвенной. Однако основной предмет геомикробиологии—это та роль, которую микроорганизмы играют а круговороте химических элементов и соединений в горных породах и подземных водах. Возникновение геомикробиологии стало возможным после открытия С. Н. Виноградским и М. Бейеринком бактерий, участвующих в определенных этапах крутоворота азота, серы, железа.
Прогресс отечественной геомикробиологии в первой половш не нашего столетия связан с именами Г. А. Надсона, Б. Л. Исаченко, Л. И. Рубенчика и ряда других ученых. В послевоенный период интенсивная работа в этом направлении началась в Институте микробиологии АН СССР. Обширный материал, полученный в ходе этих исследований, был обобщен в монографиях М. В. Иванова «Роль микробиологических процессов в генезисе месторождений самородной серы» (1964); Г. И. Каравайко, .С. И. Кузнецова, А. И. Голомзика «Роль микроорганизмов в выщелачивании металлов из руд» (1972); Г. А. Заварзина «Литотрофные микроорганизмы» (1972).
В 1972 г. Н. Н. Ляликова сообщила об открытии автотроф-Ного организма с новым типом энергетического метаболизма —Stibiobacter—бактерии, источником энергии для которой служит окисление трехвалентной сурьмы до высших окислов. В 70-е годы под руководством М. В. Иванова были проведены исследования активности бактерий в осадках озер, морей и океанов, причем для оценки активности бактерий были использованы радиоизотопные методы. Принципиальный интерес представляет собой установленный факт активного развития бактерий в глубоких слоях осадков, даже на глубине нескольких метров от поверхности ила. Широко распространены в осадках метанобразующие архебактерии и сульфатредуцирующие бактерии. Последние, как оказалось, играют весьма значительную роль в минерализации органического вещества в биосфере.
Начиная с 50-х годов исследования в области геологической микробиологии ведутся в Институте микробиологии и вирусологии АН Казахской ССР в Алма-Ате. Здесь исследовано взаимодействие бактерий с минералами, содержащими медь, цинк, свинец, мышьяк, и с ионами этих элементов, открыты автотрофные бактерии, окисляющие мышьяк. Результаты исследований обобщены в монографии А. Н. Илялетдинова «Микробиологические превращения металлов» (1984).
§ 1.
ЗЕМНОЕ ТЯГОТЕНИЕ
Все организмы, живущие на Земле, находятся под действием силы земного тяготения. Влияет ли этот фактор на развитие бактерий, остается неясным. В космических биологических исследованиях последних лет для выяснения этого вопроса предприняты специальные эксперименты. В частности, наблюдали за развитием бактерий, находящихся в условиях микрогравитации. Теоретически невесомость на микроорганизмы может влиять через перераспределение внутриклеточных части или в результате изменения взаимодействия между клеткам популяции в жидкой среде. Изменения величины массовых может оказать заметное влияние на внутриклеточные процессы только для клеток, размеры которых превышают 10 мкм. {-клетки бактерий, диаметр которых у подавляющего большинства форм меньше 10 мкм, невесомость может влиять, изменен лишь скорость их осаждения в жидкой среде. Бактерии, плотность которых несколько больше плотности воды, в водной со де находятся в состоянии, близком к невесомости. Взвешенны в водной среде бактерии все же постепенно оседают. Некоторые аэробные бактерии, выращиваемые без перемешивания, в невесомости росли лучше, видимо, благодаря меньшему оседанию на дно сосуда. Кроме того, в условиях космического полете наблюдалось некоторое увеличение частоты передачи поздним маркеров при конъюгации у кишечной палочки, т. е. образовавшиеся пары были более стабильны. Доказательств возможное прямого влияния силы гравитации на бактериальные клетки пока не получено. У бактерий не обнаружен геотаксис, т. е. способность различать верх и низ, которой обладают некоторые одноклеточные эукариоты. Вместе с тем. имеются свидетельств;
способности бактерий к определенной ориентации в пространстве об этом говорит, в частности, геометрически правильна? форма плодовых тел миксобактерий, к тому же определенные образом ориентированных относительно подлежащего субстрата.
Специфика взаимодействия бактерий с окружающей средой связана с их незначительными размерами. Бактериальная клетка только примерно в 104 раз крупнее окружающих ее молекул Она подвергается постоянной бомбардировке молекулами в результате их беспорядочного теплового движения, и эти удары ощутимы. Неравномерность пространственного распределения ударов приводит к беспорядочным перемещениям клетки каждую секунду на расстояние, примерно соответствующее ее диаметру, и к повороту примерно на 60°. Приобретаемое клеткой при этом ускорение в сотню раз превышает ее ускорениезасчёт силы земного притяжения. Таким образом, молекулярные взаимодействия, связанные с тепловым движением молекул, 1ействуют на бактерию значительно эффективнее, чем сила земного притяжения.
МАГНИТНЫЕ ПОЛЯ
Все живые организмы находятся в области магнитного поля Земли. Характер влияния магнитных полей на развитие бактерий еще мало исследован. Воздействие дополнительными более мощными полями иногда приводит к стимуляции их роста. Так. воздействие магнитным полем напряженностью в 12-Ю3 А/м приводило к некоторому ускорению роста Pseudomonas aerugi-nosa, Staphylococcus epidermidis, Halobacterium salinarium. В магнитном поле напряженностью в 24-Ю3 или 50-Ю3 А/м наблюдали их угнетение. Замедление роста Micrococcus denifrifi-cans наблюдали при 40- 104—64•104 А/м, Staphylococcus aareus и Serratia marcescens—при 120-Ю4 А/м. Действие переменных магнитных полей обычно более эффективно, чем постоянных. В естественной среде обитания бактерий магнитные поля такой напряженности не встречаются.
По всей видимости, магнитное поле Земли не оказывает заметного влияния на развитие бактерий. Однако существуют бактерии, иногда их называют магнитобактериями, способные регулировать направление своего движения в соответствии с направлением силовых линий магнитного поля. Это так называемый магнитотаксис. Магнитотаксис обнаружен у некоторых кокков и спирилл. Культуры магнитобактерий получить трудно, а их кокковидные формы в культурах пока вообще неизвестны. Сравнительно подробно исследована в культуре магнитобакте-рия quaspirillum magnetotacticum. Это микроаэрофильный хемогетеротроф.
Магнитобактерий обитают в морских маршах, заболоченных пресноводных водоемах, окислительных прудах для очистки сточных вод, т. е. в водоемах с малоподвижной водой. Их численность здесь иногда может быть весьма значительной. В области действия постоянного магнита, поднесенного к пробам воды и ила из окислительного пруда, иногда можно собрать до Ю6—Ю7 клеток таких бактерий в миллилитре воды.
Клетки магнитобактерий содержат магнитосомы—кубические или октаэдрические кристаллы магнетита. У спирилл по Длинной оси клетки расположено около 22 кристаллов с граня-ми порядка 50 нм длиной. В процессе деления дочерние клеткц получают равное количество кристаллов. Содержание железа в клетках магнитобактерий довольно велико, до 3,8%, тогда как у обычных бактерий железа содержится не более 0,025%.
В слабых магнитных полях (40—8 А/м) магнитобактерий перемещаются со скоростью до 70 мкм/с вдоль линий поля, В нашем северном полушарии бактерии плывут в сторону южного полюса магнита или северного полюса Земли. В южном полушарии, наоборот, магнитобактерий плывут в сторону южного полюса Земли. Направление силовых линий магнитного .поля Земли таково, что в результате движения вдоль этих линий бактерии в прудах и болотах оказываются у поверхности ила, где много пищи и мало молекулярного кислорода.
Воздействие сильным магнитным полем приводит к изменению направления движения магнитобактерий. Потомки этих магнитобактерий, развивающиеся в обычных условиях, т. е. под воздействием магнитного поля Земли, сохраняют направление движения, свойственное бактериям данного полушария. Если бактерии экранированы, в культуре появляются клетки, плывущие к разным полюсам, т. е. этот признак генетически не предопределен. В культуре легко могут быть получены мутанты, лишенные магнитосом и магнитотаксиса. Эти мутанты накапливаются в культуре. Не трудно представить себе, что клетки, не отягощенные столь большим количеством железа, как клетки дикого типа, получают преимущество и размножаются быстрее. Поэтому очевидно, что в природных условиях наличие магнитотаксиса для бактерий жизненно необходимо и способствует их выживанию.
Недавно обнаружено, что бактерии, обладающие магнито-таксисом, весьма многочисленны в морях. Например, в поверхностных слоях ила бассейна Санта Барбара в Тихом океане на глубине 598 м обнаружены кокки, палочки и вибрионы, содержащие магнитосомы. Магнитосомы в большом количестве накапливаются в осадках в свободном состоянии. Показано, что биогенные магнетиты обнаруживают высокий естественный остаточный магнетизм и являются его первоисточником в морских осадках.
ИЗЛУЧЕНИЯ
ВИДИМЫЙ СВЕТ
Электромагнитные волны, имеющие длину 400—740 нм, соответствуют видимому свету. Видимый свет в качестве источника энергии используют фототрофные бактерии. Цианобактерии осуществляют оксигенный фотосинтез, в процессе которого выделяется молекулярный кислород. Адаптация к разнообразным физико-химическим условиям среды, способность к азотфиксации, термофильность или галофильность некоторых форм определяют весьма широкое распространие цианобактерий на Земле. Oscillatoria Umnetica иногда способна и к аноксигенному фотосинтезу с использованием в качестве донора электронов H2S и устойчива к очень высоким Концентрациям сероводорода.
Пурпурные и зеленые бактерии осуществляют аноксигенныи фотосинтез. Он приурочен к анаэробным условиям, и большинство этих бактерий строгие анаэробы, многие нуждаются в восстановленных соединениях азота и серы. Некоторые формы способны к аэробному дыханию, но в присутствии молекулярного кислорода не могут осуществлять фотосинтез, так как уже в--низких концентрациях молекулярный кислород репрессирует синтез бактериохлорофиллов.
Развитие бактерий, осуществляющих аноксигенныи фотосинтез происходит главным образом в сильно загрязненных водоемах и гиполимнионе озер. Здесь содержатся соединения, специфически необходимые для аноксигенных фототрофов,—сероводород, молекулярный водород, простые органические вещества, образующиеся в результате брожений, которые осуществляются анаэробными бактериями, обитающими в осадках.
Пурпурные и зеленые бактерии часто развиваются в водоемах на глубине под слоем водорослей и цианобактерий, в котором происходит рост высших растений. Перечисленные фото-трофы поглощают значительную часть света в областях спектра, соответствующих адсорбционной способности их светособи-рающих антенн. Светособирающие антенны- аноксигедных фототрофов имеют другие спектральные характеристики, это позволяет им осуществлять фотосинтез за счет света, профильтрованного через клетки эукариот и цианобактерий (табл. 1).

Бактериохлорофиллы имеют максимумы поглощения в красной и инфракрасной частях спектра. Этот свет не используется для оксигенного фотосинтеза. Бактериохлорофилл б поглощает световые волны, длина которых близка к максимальной, еще допекающей их использование в процессе фотосинтеза. Кванты более длинноволнового света несут слишком мало энергии и я могут участвовать в фотохимических реакциях. Существенная особенностью бактериохлорофиллов является также то, что их включение в мембранный светособирающий комплекс приводит к смещению максимумов поглощения в красную область спектра в гораздо большей степени, чем это характерно хлорофилла а оксигенных фототрофов.
Некоторые виды пурпурных бактерий живут на мелководье под слоем влажного песка толщиной в несколько сантиметров который играет роль красного фильтра. Однако на более значительную глубину в водоем красный свет не проникает. Представители зеленых серобактерий имеют каротиноиды, улавливающие световые волны длиной около 460 нм, и способны развиваться в водоемах на глубинах, более 20 м, куда проникает свет только этой длины волны.
В конце 70-х годов были обнаружены аэробные гетеротрофные бактерии, содержащие бактериохлорофилл а. Они отнесены к нескольким родам. Представители рода Erythrobacter—мелкие грамотрицательные палочки с субполярным жгутиком. Кроме бактериохлорофилла они синтезируют каротиноидные пигменты. Ряд штаммов Erythrobacter выделен с поверхности морских макрофитов. Для Erythrobacter свет не может быть единственным источником энергии ни в аэробных, ни в -анаэробных условиях, однако используется этой бактерией в качестве дополнительного источника энергии. При освещении усиливается синтез АТФ, включение СО2 в органическое вещество клетки, наблюдается фотоингибирование дыхания, что свидетельствует о существовании у этих бактерий регуляторных механизмов энергетического обмена, позволяющих при освещении более экономно расходовать органический энергетический субстрат. При исчерпывании органического субстрата в условиях освещения жизнеспособность клеток сохраняется значительно дольше, чем в темноте, В противоположность тому, что наблюдается у пурпурных и зеленых бактерий, синтез бактериохлорофилла а у Erythrobacter стимулируется молекулярным кислородом.
Бактериохлорофилл а содержится и в мембранах гетерот рофной аэробной бактерии Protaminobacter ruber, также синтезирующей АТФ за счет энергии света. Подобная способности обааружена и у некоторых штаммов бактерий из рода Pseiidomonas.
Энергия видимого света может быть использована экстремально галофильными архебактериями, которые, однако, не являются настоящими фототрофами, поскольку не могут расти, используя свет как единственный источник энергии. Экстремально галофильная архебактерия рода Halobacterium — аэробный ге-теротроф, основное количество АТФ у которой образуется в результате процесса аэробного дыхания. Halobacterium обитает поверхностных слоях соленых озер, заливов, лиманов, где растворимость кислорода значительно ниже, чем в пресной или Юрской воде. При истощении в среде кислорода происходит репрессия генов, ответственных за синтез хромопротеина бактеоиородопсина, который включается в состав клеточной мембраны, образуя участки, обозначаемые как пурпурные мембраны. Под действием света происходит выброс протонов во внешнюю среду, создающийся при этом трансмембранный градиент концентрации протонов используется для синтеза АТФ. В тех же мембранах содержится второй пигмент—галородопсин, который при поглощении фотонов осуществляет транспорт ионов хлора в клетку, что также приводит к увеличению мембранного потенциала. Это позволяет бактериям поддерживать жизнеспособность в анаэробных условиях. Экстремально галофильные архебактерии обитают в соленых озерах, расположенных в районах с постоянной высокой инсоляцией.
Видимый свет влияет на поведение фототрофных бактерий. У (Ьототрофных эубактерий и архебактерии Halobacterium salina-rium наблюдается явление фототаксиса: бактерии способны реагировать на изменение спектрального состава света или освещенности. У эубактерий фоторецепторами служат бактерио-хлорофиллы и каротиноиды, т. е. те же пигменты, которые поглощают свет в процессе фотосинтеза. У архебактерии обнаружены специальные сенсорные пигменты. Бактериальные клетки, способные к фототаксису, резко изменяют направление движения, если попадают из более освещенного участка в менее освещенный или в совсем неосвещенный. В результате бактерии двигаются к свету—это положительный фототаксис. В некоторых условиях наблюдается отрицательный фототаксис—движение в сторону уменьшения освещенности.- Подобные реакции иногда определяют как фотофобные. При резком снижении освещенности также происходит изменение направления движения клеток, т. е. они реагируют da изменение освещенности не в пространстве, а во времени, как и в случае реакции на изменение концентрации хемоэффекторов при хемотаксисе (см. с. 126). Изменение силы света может привести к изменению скорости движения бактерий, это явление определяют как фотокинез. В реакциях фототаксиса эубактерий участвуют реакционные центры фотосинтеза. Мутанты с нарушенными реакционными центрами теряют способность к фототаксису. В этом отношении фототрофные бактерии отличаются от подвижных фототрофных эукариот, фототаксис которых не связан с фотосинтезом.
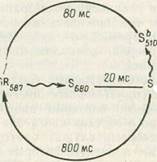
Клетки архебактерии Halobacterium salinarium проявляют фототаксис, причем красный свет действует как аттрактант, а синий—как репеллент. Оба эффекта определяются адсорбцией света одним и тем же пигментом — сенсорным родопсином (S на рис. 1). В отсутствие света сенсорный родопсин имеет максимум поглощения при 587 нм; поглощая фотон красного света, молекула переходит в состояние, при котором максимум смещается в область 680 нм и затем 373 нм. При этом генерируется: сигнал, приводящий к снижению частоты кувырканий клетки,. т. е. к аттракции. Форма 8373 может вернуться в исходное состояние за счет чисто термальной релаксации, и так фотоцикл заканчивается. Но если молекула 8373 успеет поглотить фотов синего света, она переходит в промежуточное состояние S510 а из него—в исходное. При этом генерируется сигнал, приводящий к увеличению частоты кувырканий бактерий, т. е. репеллентному эффекту. Очевидно, что в отсутствие красного света синий не вызывает этого эффекта. При освещении эффекты суммируются, и конечный результат будет зависеть от относительной интенсивности света той или иной длины волны.
Таким образом, энергия света может быть использована бактериями. Пути ее использования / прокариот значительно разнообразнее, чем у эукариот, где известен только один тип оксигенного фотосинтеза. Для некоторых бактерий, не способных использовать энергию света, он служит в качестве регулятора определенных процессов обмена. Так, у водной бактерия Pseudomonas putida наблюдали активацию светом некоторых ферментов метаболизма, что можно рассматривать как адаптацию, поскольку именно при освещении начинается фотосинтез фитопланктона, продукты которого используются гетеротрофными водными бактериями. Под фотохромностыо понимают зависимость образования-пигментов некоторыми микроорганизмами от освещения. Фотохромность свойственна миксобактерии Myxococcus xanthus особенно характерна для многих актиномицетов и близких к ним организмов. В частности, синтез каротиноидов некоторыми микобактериями стимулируется синим светом, причем процессы;
синтеза пигментов имеют светозависимые и темновые этапы.! Фотохромность может контролироваться как хромосомными, j так и плазмидными генами. Кроме того,, пигменты способны; защищать микроорганизмы от действия видимого света.
Солнечный свет может оказывать сильный антимикробный эффект. Так, более 99,9% клеток штамма Escherichia coli с нарушенными репарационными механизмами погибают после облучения солнечным светом в течение трех минут. Правда, при этом более 80% летальных повреждений связано с действием; света длиной волны менее 312 нм, действие видимого света ответственно менее чем за 1 % летальных повреждений. Видимый свет длиной волны 450 нм индуцирует замены пар оснований д мутации сдвига рамки у Е. coli. Световые волны длиной 550 им и особенно 410 нм вызывают фотолизис клеток Myxococcus xanthus. Эффект определяется поглощением света желе-зопорфиринами.
Существуют вещества фотосенсибилизаторы, в- молекуле ко-тооых имеется хромофор, поглощающий свет и передающий его энергию другим молекулам, не способным поглощать свет. Через бесцветные клетки свет проходит без последствий, но если в такую клетку введен фотосенсибилизатор, она повреждается. Известно более 400 веществ, обладающих свойствами фотосенсибилизаторов. Их хромофорами обычно оказываются циклические ядра. Фотосенсибилизаторы содержатся в промышленных стоках, смоге, многие лекарства представляют собой фотосенсибилизаторы, в том числе сульфаниламиды, ряд антибиотиков, хинин, рибофлавин, метиленовый синий и другие красители. Среди .природных веществ фотосенсибилизаторами являются псоралены (фурокумарины), хлорофилл, фикобилины, порфири-ны и др. Многие растения содержат псоралены, свет в их присутствии повреждает ДНК. Порфирины находятся во всех аэробных и некоторых анаэробных клетках. При нарушениях в путях метаболизма могут накапливаться промежуточные продукты синтеза порфиринов, также действующие как фотосенсибилизаторы.
Некоторые фотосенсибилизаторы действуют только в присутствии кислорода—это так называемый фотодинамический эффект (см. с. 92).
Дата добавления: 2020-10-01; просмотров: 994;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
