Доантигенный этап развития Т-лимфоцитов
Доантигенный (первичный) Т-лимфоцитопоэз осуществляется в корковом веществе тимуса из пре-Т-клеток или малодифференцированных лимфобластов, мигрирующих из красного костного мозга. При этом в процессе превращения пре-Т-клетки в наивный специфичный Т-лимфоцит, наряду с митотическими делениями, происходит и изменение экспрессии фенотипических Т-клеточных маркеров. В частности, первые мигранты из красного костного мозга – пре-Т-клетки – представляют собой малодифференцированные лимфобласты, имеющие определенный набор поверхностных молекул, но лишенные основных поверхностных маркеров дифференцировки Т-лимфоцитов: СD4 и CD8-корецепторов, типичных соотвественно для Т-хелперов/Т-клеток воспаления и Т-киллеров. В связи с отсутствием на поверхности малодифференцированных лимфобластов специфических рецепторов Т-лимфоцитов (СD4 и CD8) их называют «двойными негативами». Эти клетки заселяют верхнюю часть коры тимуса – субкапсулярную его зону и в полностью сформированном тимусе составляют незначительный его пул (5% от общего количества тимоцитов).
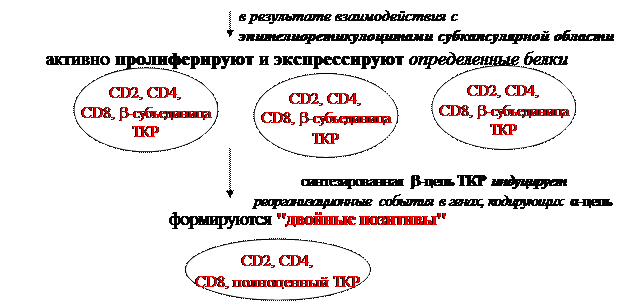
Взаимодействие малодифференцированного лимфобласта с эпителиоретикулоцитами субкапсулярной области индуцирует экспрессию в нем первого специфического маркера Т-клеток: СD2, который обладает адгезивными свойствами и относится к суперсемейству иммунноглобулинов. Лимфобласты субкапсулярной зоны тимуса, находясь в тесном контакте с эпителиоретикулоцитами, от которых получают питательные вещества и микроэлементы, активно пролиферируют и завершают свой путь развития в данной зоне умеренной экспрессией СD4, CD8-молекул,а также b-субъединиц специфического Т-клеточного рецептора (ТКР), после чего перемещаются в более глубокие слои коркового вещества тимуса. При этом в субкапсулярной зоне в результате взаимодействия пролиферирующих лимфобластов с эпителиоретикулоцитами происходит первое реорганизационное событие в исходном наборе генов, кодирующих b-цепь ТКР –один из 2-х D-генных сегментов, кодирующих вариабельный домен b-цепи, случайным образом объединяется с одним из 12-ти J-сегментов, в результате чего образуется информационный DJ-локус. После этого на уровне субкупсулярной зоны в развивающихся Т-лимфоцитах происходит второе реорганизационное событие – объединение одного из 30 V-генов с сформированным DJ-локусом, что приводит к формированию полноценного информационного VDJ-локуса, кодирующего вариабельный домен b-цепи ТКР.На этом этапе развития Т-лимфоцитов синтезируемая b-цепь ТКР еще остается в цитоплазме.
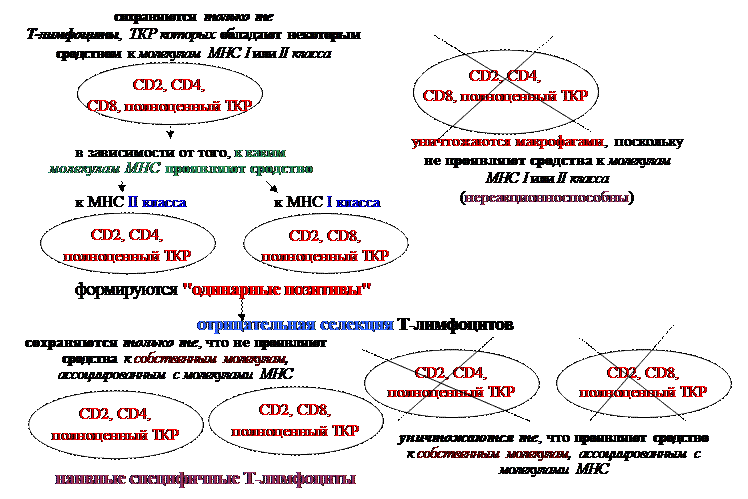
В результате взаимодействия таких Т-лимфоцитов, мигрирующих из субкапсулярной зоны, с эпителиоретикулоцитами коры происходит активная одновременная экспрессия на их поверхности корецепторов СD4, CD8, а также полноценного ТКР,состоящего изα- и b-цепей, в связи с чем Т-лимфоциты на данной стадии дифференцировки называют «двойными позитивами». При этом в качестве сигнала, индуцирующего реорганизационные события в генах, кодирующих α-цепь(образование информационного VJ-локуса), а, следовательно, образование полноценного ТКР, а также выход на клеточную поверхность СD4 и CD8-рецепторов, служит синтезированная b-цепь ТКР.Взаимодействие же развивающихся Т-лимфоцитов с ретикулоэпителиальными клетками глубоких слоев коркового вещества тимуса сопровождается положительной селекцией Т-лимфоцитов, приводящей к сохранению только тех Т-лимфоцитов, ТКР которых обладают некоторым сродством к молекулам МНС I или II класса, представленным на поверхности ретикулоэпителиоцитов. В частности, на стадии умеренной экспрессии ТКР Т-лимоциты представляют собой сырой материал для отбора. В коре тимуса, содержащей ретикулоэпителиальные клетки, экспрессирующие молекулы МНС как I, так и II класса, происходят основные селекционные события. Сущность их заключается в том, что если Т-лимфоциты обладают некоторым сродством к молекулам МНС I или II класса, то они подвергаются дальнейшей дифференцировке. Развитие же Т-лимфоцитов, ТКР которых вообще не соотвествует специфичностям молекул МНС, завершается в тимусе. Но при этом, если клетка не выдержала первичного положительного отбора, она еще некоторое время (3-4 суток) сохраняет жизнеспособность и продолжает реорганизовывать гены α-цепи, представляя несколько иные, чем исходно, по специфичности ТКР. Иными словами, Т-лимфоцит путем изменения пространственной структуры α-цепей пытается подогнать структуру своих ТКРтаким образом, чтобы иметьвозможность вступать во взаимодействие с антигенсвязывающим участком МНС, который служит для представления антигенного пептида. Без этого дополнительного механизма реорганизации генов α-цепи гибель Т-лимфоцитов в результате положительной селекции в тимусе была еще более высокой по сравнению с существующей (в норме погибает до 95% от образующихся Т-лимфоцитов на стадии положительной селекции).
Определяющую роль в положительной селекции Т-лимфоцитов играет степень сродства ТКР Т-лимфоцитак молекуле МНС. Если это сродство очень велико, то такой Т-лимфоцитподвергаетсяапоптозу. В том же случае, если это сродство Т-лимфоцита к собственным молекулам МНС существует, но относительно не велико, он сохраняется в результате положительной селекции. Если же Т-лимфоцит вообще не проявляет сродства к какому-то из классов МНС, то он уничтожается в результате положительной селекции, поскольку будет нереакционноспособен.
Вместе с тем, на этапе формирования клонов Т-лимфоцитов, способных взаимодействовать с одной из молекул МНС, процесс их дифференцировки не заканчивается. Т-лимфоциты, прошедшие положительную селекцию, первоначально экспрессируют как СD4-, так и CD8-корецепторы. Но как только отбор на специфичность произошел, Т-лимфоцит теряет один из корецепторов. В частности, если ТКР Т-лимфоцита вывявил специфичность к молекулам МНС I класса, то сохраняется корецептор CD8, но утрачивается CD4 (т.е. дифференцируется предшественник Т-киллера). В случае, если в результате положительной селекции выявилось сродство ТКР Т-лимфоцита к молекулам МНС II класса, то в них продолжается экспрессия CD4, но прекращается экспрессия CD8 (клетка дифференцируется в предшественник Т-хелпера или Т-клетки воспаления). Таким образом, по мере дальнейшего доантигенного созревания Т-клетки после первичной селекции постепенно (в нижних участках коркового вещества, расположенных на границе с мозговым, в т.н. корково-медуллярной зоне) утрачивают один из видов корецепторов СD4 или CD8 (превращаются из «двойных позитивов» в «одинарные позитивы»), в результате чего появляются наивные специфичные Т-лимфоциты двух основных субпопуляций: СD4-ТКР-клетки (Т-хелперы или Т-клетки воспаления) и CD8-ТКР (Т-киллеры), часть которых мигрирует в мозговое вещество тимуса, а часть – в периферические лимфоидные органы. Дальнейшая антигензависимая дифференцировка СD4-Т-лимфоцитов в Т-хелперы или Т-клетки воспаления будет зависеть от того, с комплексом «антигенная детерминанта-молекула МНС II класса» какой клетки (В-лимфоцита или макрофага) провзаимодействует СD4-Т-лимфоцит. В случае взаимодействия с антигенным комплексом, представленным на поверхности В-лимфоцита, СD4- клетка
Основные этапы доантигенного развития Т-лимфоцитов
Исходная стадия
субкапсулярная зона коркового вещества тимуса
 |
малодифференцированные лимфобласты, мигрирующие из красного костного мозга
(ЛИШЕНЫ основных поверхностных маркеров дифференцировки Т-лимфоцитов:
СD4 и CD8-корецепторов, «двойные негативы»)
 | |||||||
| |||||||
 | |||||||
 |
превратится в Т-хелпер, а при взаимодействии с антигенным комплексом, представленным на поверхности макрофага, – в Т-клетку воспаления.
Кроме того, в корково-медуллярной зоне на стадии «двойных позитивов» происходит и отрицательная селекция Т-лимфоцитов,заключающаяся в элиминации Т-лимфоцитов проявляющих сродство к собственным молекулам, ассоциированным с молекулами МНС. Фенотип Т-лимфоцитов, не прошедших отбор на специфичность (положительную или отрицательную селекцию), соотвествует двойным позитивам (СD4CD8) и указывает на незавершенность дифференцировки.
Таким образом, Т-лимфоциты, погибающие в тимусе, не выдерживают два условия положительной селекции (проявляют либо очень высокую аффинность ТКР к молекулам МНС, либо вообще ее не проявляют), либо проявляют реакции на собственные антигены, в результате чего элиминируются отрицательной селекцией. Меньшая же часть популяции Т-лимфоцитов, прошедших жесткие условия отбора на специфичность, покидает тимус и расселяется в периферических органах иммунной системы. Весь путь доантигенного развития Т-лимфоцитов создает потенциал для возможной в будущем (в периферических органах иммунной системы) встречи с различными чужеродными антигенами, но при этом исключает выход в циркуляции наивных Т-лимфоцитов, настроенных на собственные антигены и тех клеток, которые не проявляют сродства к молекулам МНС, в результате чего окажутся неспособными взаимодействовать с комплексами «антигенная детерминанта-молекула МНС» на поверхности антигенпрезентирующих клеток, а, значит, и неспособными пройти антигензависимую дифференцировку, превратится в зрелые и развить иммунный ответ.
Дата добавления: 2016-07-18; просмотров: 2893;
Поиск по сайту
Узнать еще
- Filariidae:. Onchocerca volvulus Систематика, морфология, цикл развития, патогенное действие, диагностика и профилактика онхоцеркоза и лоаоза.
- I этап –главные идеи:
- II этап – анаэробный (бескислородный)
- II. Завоевание Китая маньчжурами. Экономическое положение страны в XVII – начале XIX вв.: аграрная политика Цинской династии, особенности развития городского ремесла
- II. Особенности развития турецкой буржуазии. Становление младотурецкого движения
- III Всебелорусское собрание. Программа социально-экономического развития Республики Беларусь на 2006 – 2010 гг.
- III. ТРЕТИЙ ЭТАП ОБУЧЕНИЯ
- IV этап – обособление тела зародыша

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
