ТРАНСМЕМБРАННЫЙ ПОТЕНЦИАЛ ДЕЙСТВИЯ
Акционный трансмембранный потенциал возникает при механическом, электрическом, химическом или иного рода раздражении клетки. Перечисленные раздражители снижают потенциал покоя до критической величины порогового потенциала, которая для большинства сердечных клеток равна примерно 60 мв. По достижении порогового потенциала наступает быстрая активация, деполяризация клетки (фаза 0), а затем постепенно происходит реполяризация клетки (фазы 1, 2, 3). Акционный потенциал включает в себя деполяризацию и реполяризацию клетки. Это значит, что он состоит из фазы 0, 1, 2 и 3 трансмембранного потенциала. Кривой акционного потенциала свойственны следующие особенности: деполяризация (фаза 0) представлена как круто поднимающаяся вверх почти вертикальная прямая линия, которая переходит над нулевым уровнем и достигает примерно +30 мв; фаза 1 реполяризации — короткая, круто спускающаяся вниз кривая; фаза 2 реполяризации имеет форму плато и находится около уровня нулевого потенциала, а фаза 3 представляет собой круто спускающуюся вниз линию, заканчивающуюся у уровня потенциала покоя (фаза 4). Ионные механизмы отдельных фаз акционного потенциала еще не вполне уточнены.

Трансмембранный электрический потенциал клетки сократительного миокарда; электрокардиограмма поверхности тела и трансмембранное движение катионов.
Наиболее важным моментом активирования клетки является достижение порогового потенциала. Тогда проницаемость клеточной мембраны для ионов натрия резко повышается и они быстро поступают в клетку. Такое движение ионов натрия снаружи через клеточную мембрану внутрь клетки, известное как „ток Na+", вызывает электрическую ак-тивицию сердечной клетки — деполяризацию, фазу 0 потенциала действия. Во время деполяризации внутренняя среда клетки быстро становится электроположительной в отношении внеклеточной жидкости. Имеются данные, указывающие на то, что фаза 1 реполяризации обусловливается поступлением ионов хлора в клеткy (Trautwein), а фаза 2 реполяризации является результатом сравнительно более низкого по степени и скорости поступления ионов натрия в клетку, а в некоторых клетках наблюдается и переход ионов кальция в клетку (Reuter). В фазе 3 реполяризации начинается выход ионов калия изнутри наружу через клеточную мембрану, известный как „ток К+" (McAllister. Noble).
Электрокардиограмма, регистрируемая с поверхности тела, представляет собой суммарный эффект (интегральное изображение) трансмембранных потенциалов активированных сердечных клеток. Волна Р электрограммы представляет собой выражение предсердной, а комплекс QRS — желудочковой деполяризации. Сегмент ST и волна Т — выражение желудочковой реполяризации. Точнее говоря, комплекс QRS электрокардиограммы соответствует фазе 0 трансмембранного потенциала, сегмент Т—фазе 1 и фазе 2, а волна Т—фазе 3.
Существуют четыре главных электрофизнологических свойства сердца: автоматизм, проводимость, возбудимость и рефрактерность.
АВТОМАТИЗМ (Механизм зарождения импульсов)
Сердечная мышца состоит из двух видов клеток — клеток проводниковой системы и клеток сократительного миокарда.Только некоторым клеткам проводниковой системы присущи автоматические свойства — синоаурикулярному узлу,специализированным проводниковым путям в предсердиях, клеткам септальных створок атриовентрикулярных клапанов, нижней части пучка Гиса, обеим его ножкам и волокнам сети Пуркииье. Автоматизм — это способность спонтанно активироваться, т. е. создавать электрические импульсы без помощи нервной стимуляции. Клетки проводниковой системы, обладающие этим свойством, называются также пейсмекерными клетками, т. е. клетками, которые являются или могут стать электрическим водителем ритма сердца. Механизм автоматизма можно понять, сравнивая трансмембранные потенциалы клетки проводниковой системы и клетки сократительного миокарда. Наиболее существенная разница между ними состоит в диастолической фазе, т. е. у них различный трансмембранный потенциал в состоянии покоя (фаза 4). Трансмембранный потенциал клеток сократительного миокарда остается постоянным в период диастолы, и на трансмембранной электрограмме видна прямая горизонтальная линия. Диастолический потенциал клеток проводниковой системы изменяется, медленно, спонтанно понижаясь до критической величины порогового потенциала, после чего наступает быстрая активация клетки. Электрограмма трансмембранного потенциала показывает полого, с небольшим наклоном, медленно поднимающуюся кверху прямую диастолическую линию. Этот процесс, называемый спонтанной диастолической деполяризацией, представляет собой механизм зарождения импульсов, автоматизм клеток водителя ритма. Изменения ионов, обусловливающие спонтанную диастолическую деполяризацию и приводящие к спонтанному созданию электрических импульсов, еще не выяснены окончательно. Основное значение имеют особенности проницаемости мембраны и „ток К+" в диастолический период клеток водителя ритма; непосредственно после окончания реполяризации проницаемость клеточной мембраны для нона калия оказывается значительной и поэтому большое количество ионов калия выходит из клетки (Noble, Tsien). Во время диастолы проницаемость клеточной мембраны для ионов калия, как и выведение ионов калия из клетки постепенно уменьшаются. Вместе с тем, налицо некоторая проницаемость для ионов натрия и небольшое количество этих ионов проникает в клетку. Величина этого тока ионов натрия сохраняется почти на одном уровне в течение всей диастолы (Weidmann). Оба процесса — постепенно уменьшающееся выведение ионов калия и постепенное поступление в небольшом количестве ионов натрия — приводят к медленному увеличению положительно заряженных ионов в клетке. Таким образом наступает постепенное уменьшение отрицательного значения диастолического трансмембранного потенциала. Когда диастолический трансмембранный потенциал достигнет величины порогового, т.е. когда он снизится с исходной величины—90 мв, до пороговой— 60 мв, тогда проницаемость клеточной мембраны для ионов натрия резко повышается и большие количества его быстро поступают в клетку. Такой внезапный поток ионов натрия вызывает возникновение электрического импульса (деполяризация, фаза 0), представляющего собой сердечный импульс возбуждения. Последний, следовательно, вызывается вследствие движения ионов натрия снаружи через клеточные мембраны внутрь клеток проводниковой системы. По сути дела, речь идет о цепной электрофизиологической реакции. при которой небольшие воздействия тока, возникшие спонтанно из-за пониженного выхода ионов калия и незначительного поступления ионов натрия во время диастолы, резко повышают проницаемость клеточной мембраны для ионов натрия, и последние своим движением вызывают импульс возбуждения. Сердечный импульс возбуждения, ио своей сущности, является электрическим импульсом. В основе сердечного автоматизма лежат ионообменные процессы в клетке.
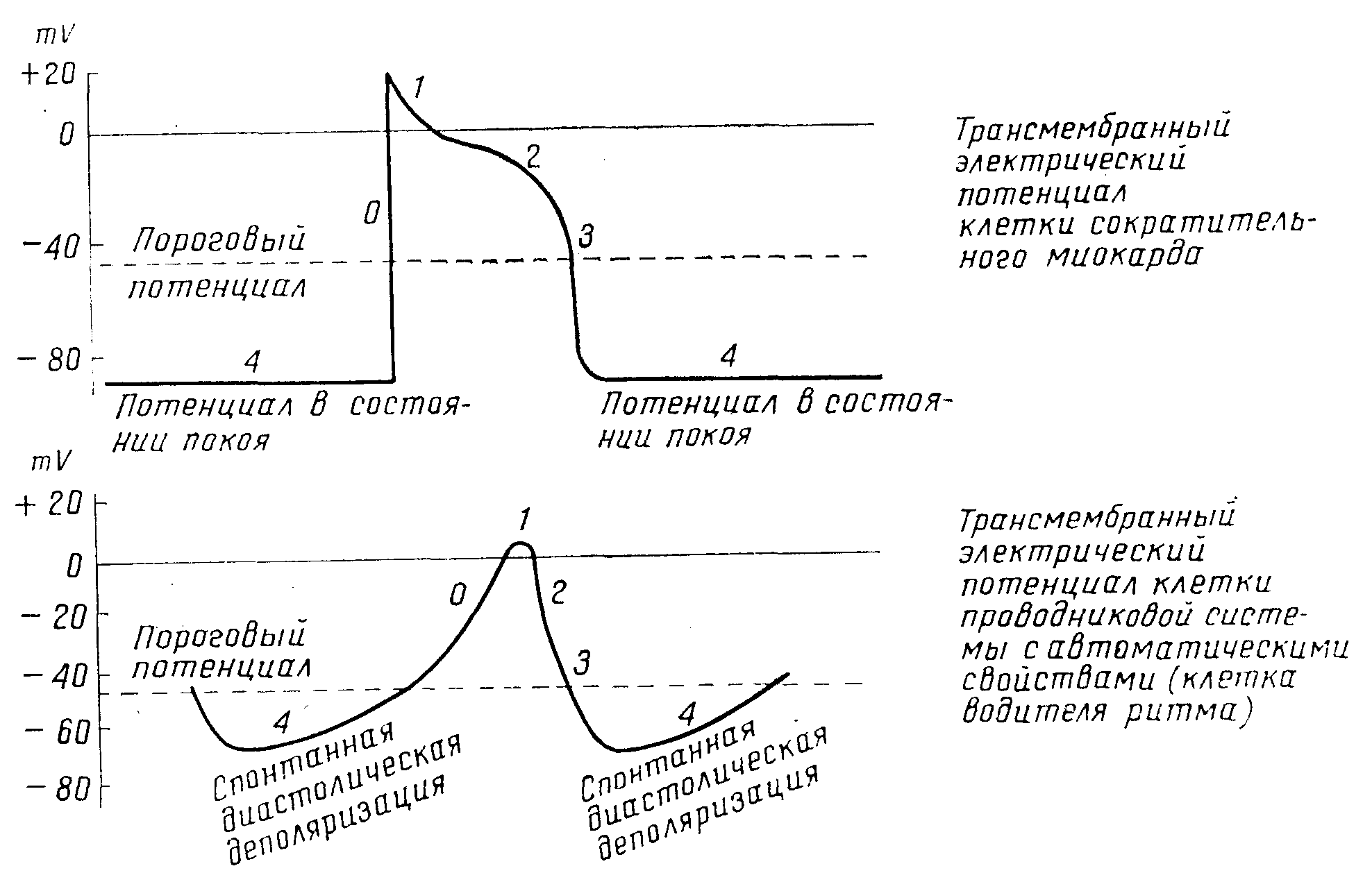
Трансмембранные потенциалы клетки сократительного миокарда и клетки проводниковой системы — автоматическими свойствами. Наиболее существенная разница между ними заключается в диастолической фазе (фаза 4).
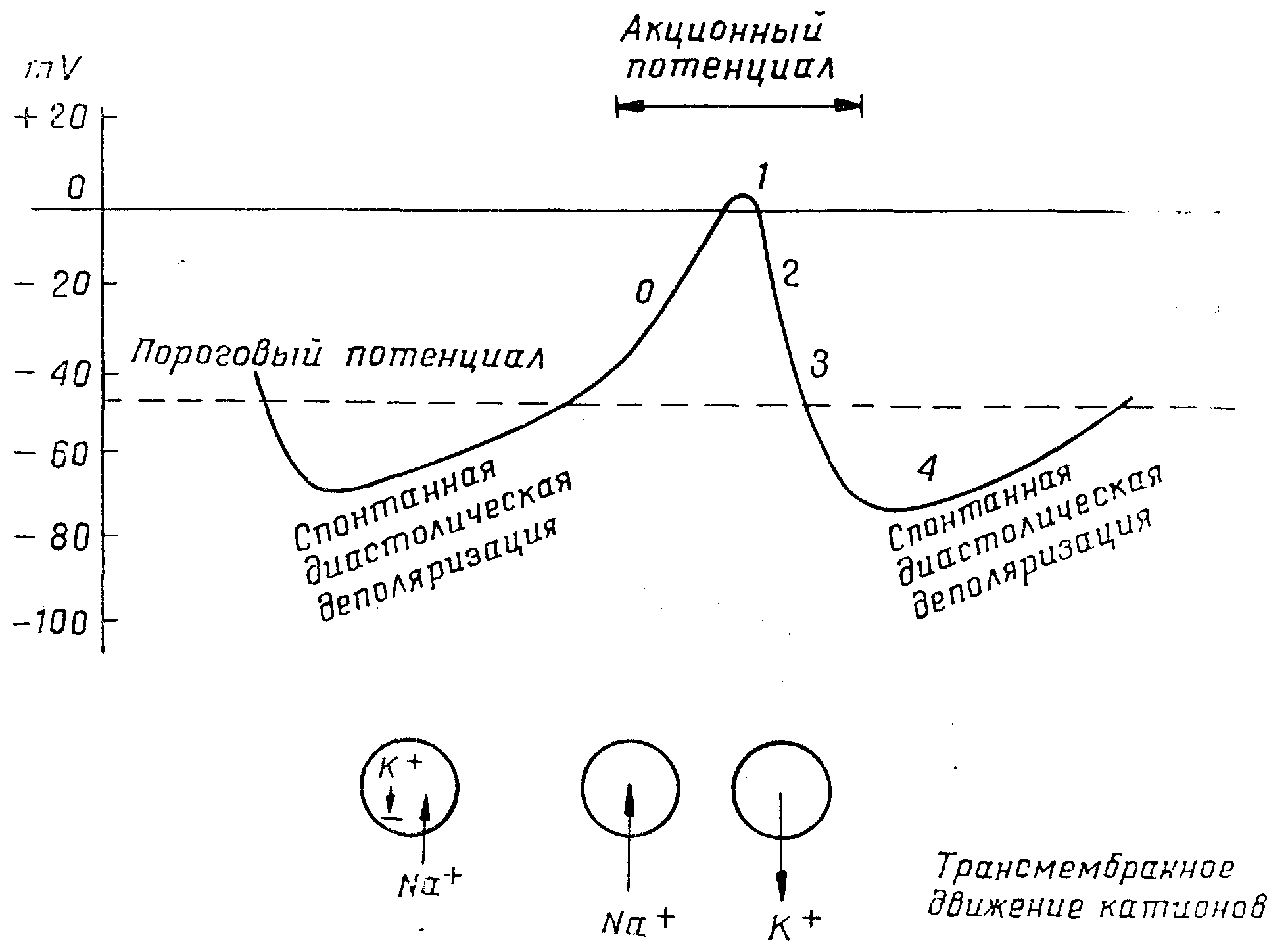
Трансмембранный потенциал клетки, проводниковой системы с автоматическими свойствами (клетка водителя ритма). Автоматизм является результатом спонтанной диастолической деполяризации. Когда степень спонтанной диастолической деполяризации достигает порогового потенциала, наступает электрическое активирование клетки (акционный потенциал). Показаны движения катионов через клеточную мембрану.
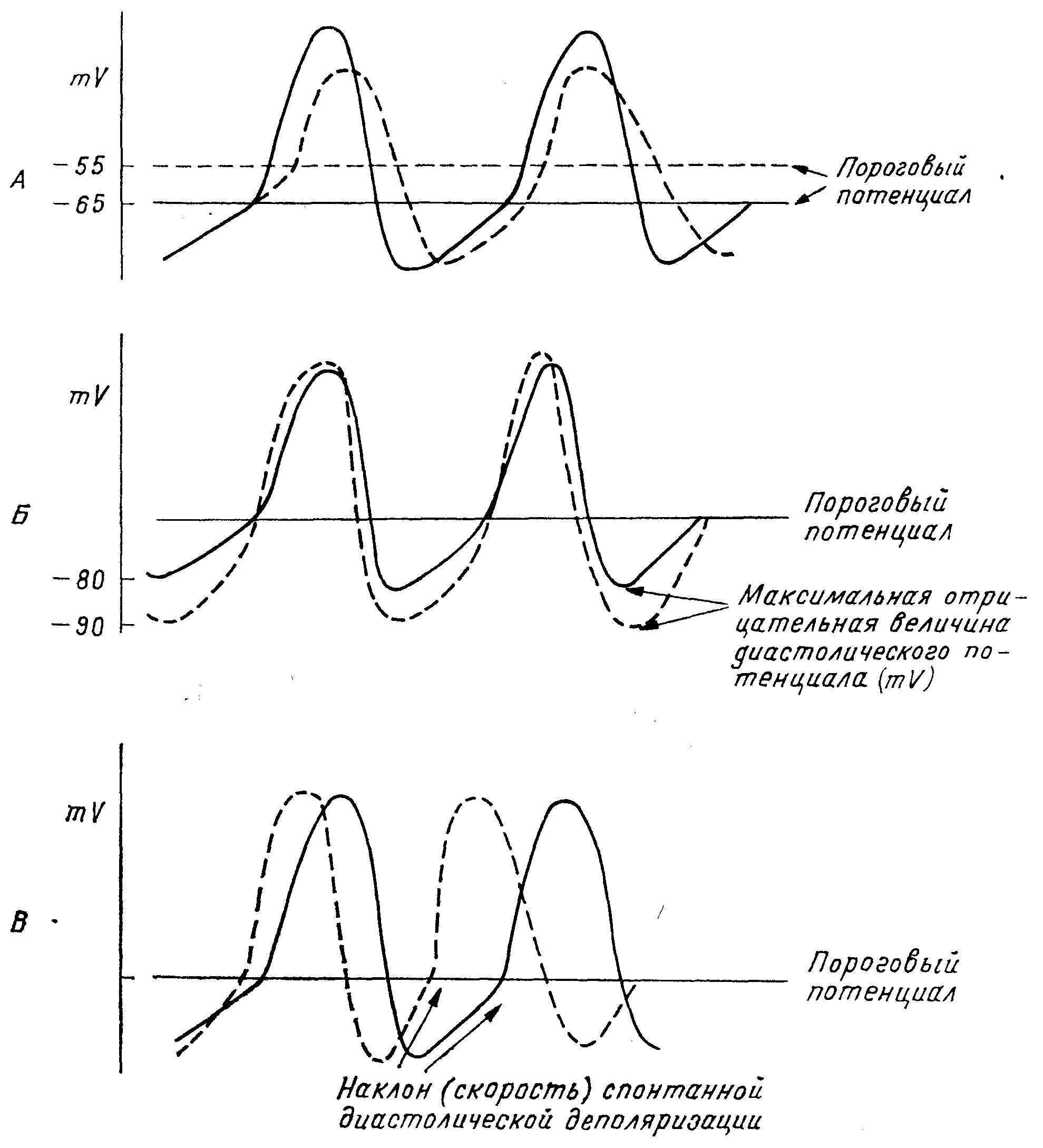
Параметры трансмембранного потенциала, определяющие степень автоматизма сердца.
А — изменения величины порогового потенциала. Б — изменения величины максимального диастолического потенциал|
в начале диастолического периода (Vm). В — Изменение наклона (скорости) спонтанной диастолической деполяризации.
Степень выраженности сердечного автоматизма зависит от четырех главных параметров трансмембранного потенциала (рис. 7):
1. Величины порогового потенциала. Чем больше, чем более отрицателен пороговый потенциал, тем более выраженными будут автоматические свойства клетки и, наоборот. Так, при пороговом потенциале —65 мв наблюдается более сильно выраженный автоматизм, чем при пороговом потенциале —55 мв.
2. Максимальной отрицательной величины диастолического потенциала (V). Это величина в начале диастолического периода, непосредственно после окончания реполяризации. Чем выше максимальная величина диастолического потенциала, тем более выражены будут автоматические свойства клетки и, наоборот. Так, при максимальном диастолическом потенциале —90 мв автоматизм будет более сильным, чем при величине того же потенциала —80 мв.
3. Разности между максимальным диастолическим и пороговым потенциалами. Чем меньше эта разность, тем быстрее спонтанная диастолическая реполяризация достигает порогового потенциала и тем более сильно выраженным будет автоматизм и, наоборот.
4. Наклона (быстроты, скорости) спонтанной диастолической деполяризации. Чем быстрее происходит спонтанная диастолическая деполяризация и чем больше ее наклон, тем быстрее будут создаваться автоматические электрические импульсы и, наоборот.
В нормальных условиях синусовый узел определяет ритм всего сердца ввиду того, что описанные выше электрофизиологические процессы спонтанной диастолической деполяризации быстрее и интенсивнее всего протекают в его клетках (рис. 8). Чем ниже спускаемся по проводниковой системе, тем слабее автоматизм клеток водителя ритма. Обычно частота синусовых импульсов — 60—100, атриовентрикулярного узла — 40—60, а проводниковой системы в желудочках — менее 40 в минуту. Клетки водителя ритма в предсердиях, атриовентикулярном узле и пучке Гиса называются автоматическими центрами второго порядка (наджелудочковыми), а эти клетки в обеих ножках пучка Гиса и сети Пуркинье — автоматическими центрами третьего порядка (желудочковыми). В том случае, когда синусовый узел временно подавлен или же функции его полностью выпали, водителем ритма становится автоматический центр второго, а затем третьего порядка. Нормально импульсы синусового узла подавляют активность нижележащих автоматических центров.

Конфигурация кривой акционного потенциала различных типов клеток. Продолжительность акционного потенциала прогрессивно нарастает от предсердий к клеткам Пуркинье. Спонтанная диастолическая деполяризация (фаза 4) быстрее всего и интенсивнее всего протекает в клетках синоаурикулярного узла, вследствие чего нормальный синусовый узел определяет ритм всего сердца.
ПРОВОДИМОСТЬ
Проводимость является свойством всех клеток миокарда проводить импульсы возбуждения к окружающим их соседним клеткам (рис. 9). Проведение представляет собой последовательное развертывание описанной выше электрофизиологической реакции.
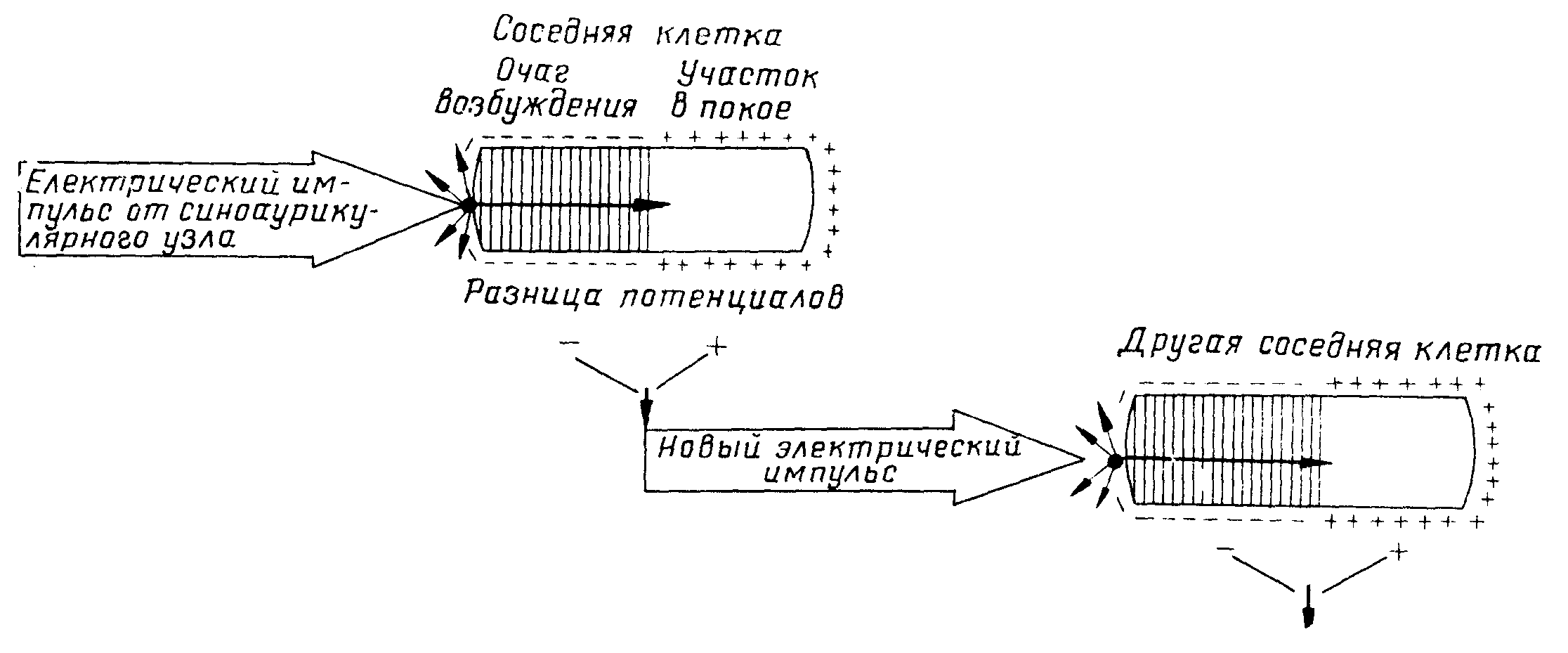
Схема механизма проводимости в сердце
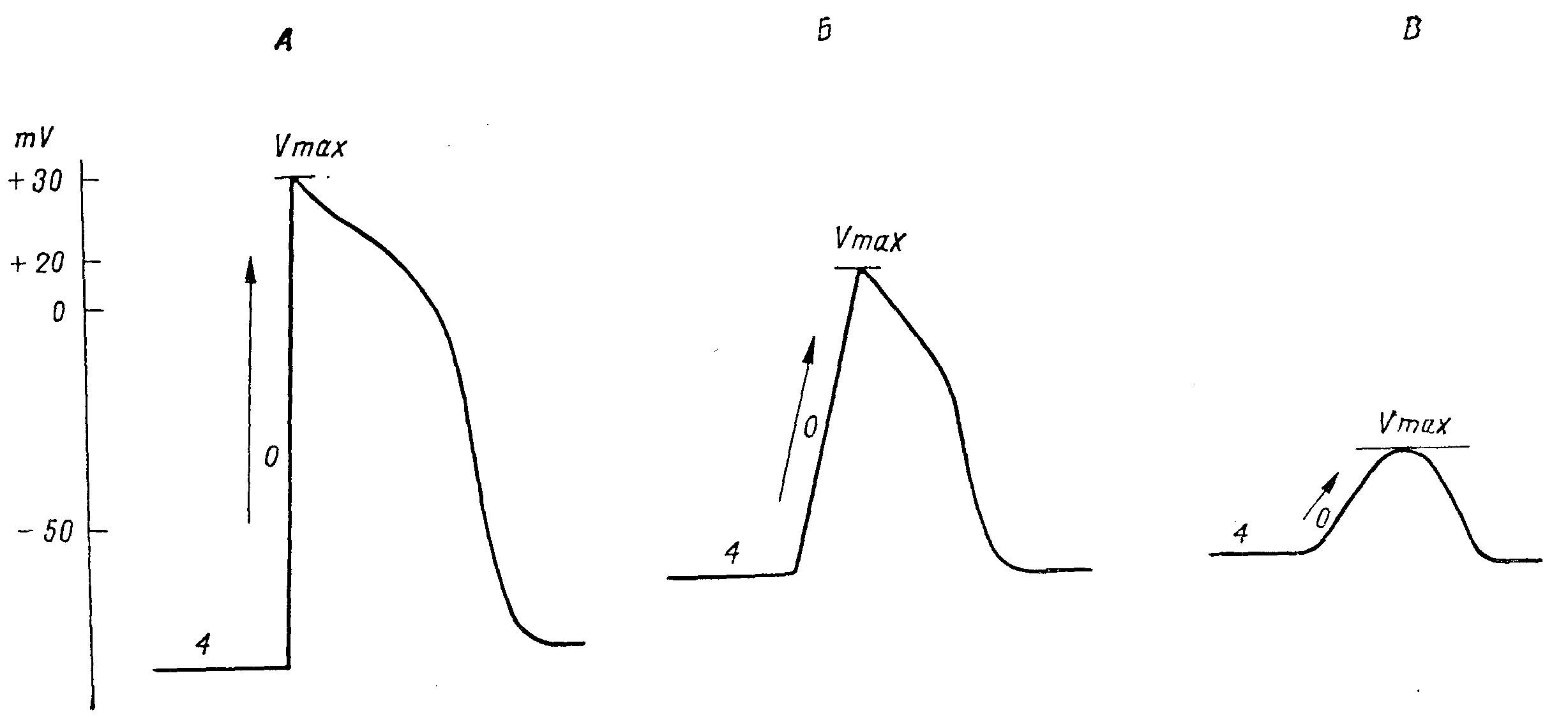
Параметры трансмембранного потенциала, определяющие скорость проведения. Представлены три клетки — Л, Б, В — с различной скоростью проведения. Проводимость клетки Л самая быстрая, так как максимальная амплитуда акционного потенциала (Vmax) большая (+30 мв); деполяризация (фаза 0) происходит быстро и круто, и потенциал в покое перед моментом стимуляции сильно отрицательный. Проводимость клетки В самая медленная, так как максимальная амплитуда акционного потенциала (Vmax) очень мала, деполяризация — (фаза 0) — происходит медленно, и потенциал в покое (фаза 4) перед моментом стимуляции имеет небольшое отрицательное значение. По скорости проведения клетк Б занимает среднее место.
Поступающие из синусового узла или из другого нижележащего автоматического центра электрические импульсы воздействуют на мембраны клеток, расположенных около него, и вызывают перемещение ионов, о котором шла речь выше. По достижении порогового потенциала соседних клеток наступает быстрое движение ионов натрия внутрь их. Это движение ионов натрия отражается в электрическом токе (потенциале действия), который действует как деполяризующий стимул на другие соседние клетки и, таким образом, электрический ток проводится из клетки в клетку и постепенно проходит через все сердце. Цитоплазма сердечных клеток и межклеточная сердечная жидкость обладают небольшим электрическим сопротивлением и являются хорошими проводниками электрического тока. Сначала активируется один конец клеток. Между возбужденным, электроотрицательным и находящимся в покое электроположительным участком клеток создается разность потенциалов. Через клетку проходит ток, воздействующий на другую группу соседних клеток, и таким образом электрическое возбуждение распространяется вперед.
Скорость проведения зависит от следующих параметров трансмембранного потенциала (рис. 10):
1. Максимальной амплитуды потенциала действия (Vmax). Чем больше максимальная величина акционного потенциала, тем быстрее будет происходить проведение и, наоборот. Например, при максимальной амплитуде акционного тока 4-30 мв проведение будет быстрее, чем при величине его 4-20 мв.
2. Скорости деполяризации фазы 0 (Vmax).Чем быстрее и круче поднимается фаза 0 трансмембранного потенциала, тем больше будет скорость проведения и, наоборот.
3. Амплитуды диастолического потенциала в момент стимуляции. Чем больше (более отрицателен) потенциал в состоянии покоя перед моментом стимуляции клетки, тем быстрее происходит проведение и, наоборот. Когда диастолический потенциал сильно снижен — до 50 мв или больше, в момент перед активацией не создается потенциал действия, Vmax будет иметь нулевую величину, т. е. произойдет блокада проведения.
4. Величины порогового потенциала и разницы между диастолическим и пороговым потенциалом. Чем выше уровень порогового потенциала и больше разница между диастолическим и пороговым потенциалом в момент стимуляции, тем медленнее будет проводиться импульс.
Скорость проведения электрического импульса зависит от строения и функциональных особенностей разных отделов проводниковой системы и миокарда. Скорость проведения наиболее высокая в клетках Пуркинье (400мм веек.) и наиболее низкая в атриовентрикулярном узле (200 мм в сек.). Проводимость пучка Гиса — 800—1000 мм в сек., сократительной мускулатуры желудочков — 400 мм в сек.
ВОЗБУДИМОСТЬ
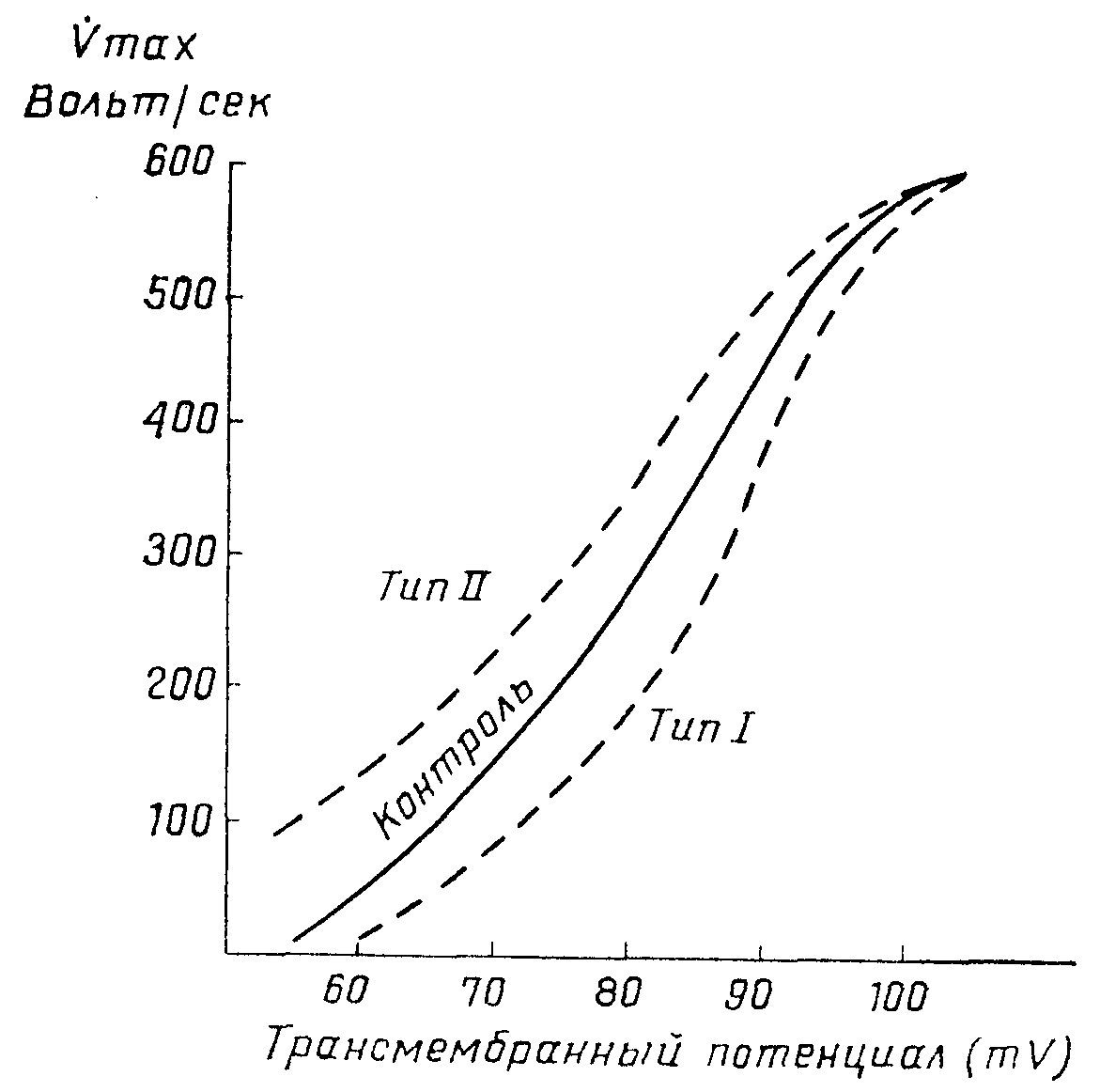
Характеристика степени возбудимости клеток. Максимальная скорость деполяризации (Vmax) зависит от трансмембранного потенциала в покое (mV) до момента раздражения. Чем больше (более негативно) значение мембранного потенциала перед раздражением, тем больше максимальная скорость деполяризации, т. е. тем больше возбудимость клетки.
Это свойство живой ткани реагировать на различные раздражители. Возбудимостью обладают клетки проводниковой системы и сократительные клетки сердца. Возбудимость сердечной клетки выражается в ее способности активироваться электрически, т. е. генерировать потенциал действия в результате разного рода раздражении. Если бы способность возбуждаться не была свойственна всем сердечным клеткам, невозможным было бы осуществление тотального электрического активирования всего сердца, независимо от того, что в некоторых клетках сердца может существовать автоматизм. Благодаря возбудимости возможно распространение электрического сердечного импульса. Возбудимость следует различать от автоматизма, являющегося свойством некоторых сердечных клеток спонтанно создавать электрические импульсы. Оба эти свойства сердца изменяются параллельно при различных состояниях, однако из этого правила бывают и исключения. Так, например, при интоксикации препаратами наперстянки можно наблюдать повышенный автоматизм, но пониженную возбудимость. Возбудимость сердца тем выше, чем более слабый раздражитель в состоянии вызвать электрическую активность клетки. Возбудимость сердца подчиняется закону „все или ничего". Это значит, что подпороговые раздражители не вызывают акционного потенциала, тогда как раздражители, имеющие пороговую величину, вызывают максимальный по силе и скорости акционный потенциал. Повышение силы раздражения не ведет к повышению степени акционного потенциала. Одной из наиболее важных закономерностей клеточной электрифизиологии сердца, характеризующей степень клеточной возбудимости, является так называемый ответ клеточной мембраны на раздражение (Weidemann, Hoffman и сотр.). Ответ клеточной мембраны на раздражение определяется как отношение максимальной скорости фазы 0, т. е. деполяризации (Vmax), к величине мембранного потенциала перед моментом возбуждения (рис. 11). Максимальная скорость деполяризации является функцией мембранного потенциала непосредственно перед раздражением. Чем выше (более отрицательна) величина мембранного потенциала перед раздражением, тем больше будет мак-мимальная скорость деполяризации, т. е. тем сильнее будет возбудимость клетки.
РЕФРАКТЕРНОСТЬ
Возбудимость сердечной клетки изменяется в отдельные периоды сердечного цикла. Во время систолы сердечная клетка не возбуждается, т. е. она рефрактерна к раздражению. Во время диастолы возбудимость сердечной клетки восстанавливается. Рефрактерность—это невозможность активизированной сердечной клетки снова] активироваться при дополнительном раздражении. Сердечная клетка, охваченная процессом электрического возбуждения и обладающая акционным потенциалом, не может создать другое дополнительное электрическое возбуждение, другой акционный потенциал. Электрическое возбуждение полностью вовлекает в процесс систему ионов натрия клетки, вследствие чего отсутствует ионный субстрат, который мог бы ответить на дополнительное раздражение.
Различают три степени рефрактерности, соотв. периода: абсолютный, эффективный и относительный (релятивный) рефрактерный период (рис. 12).
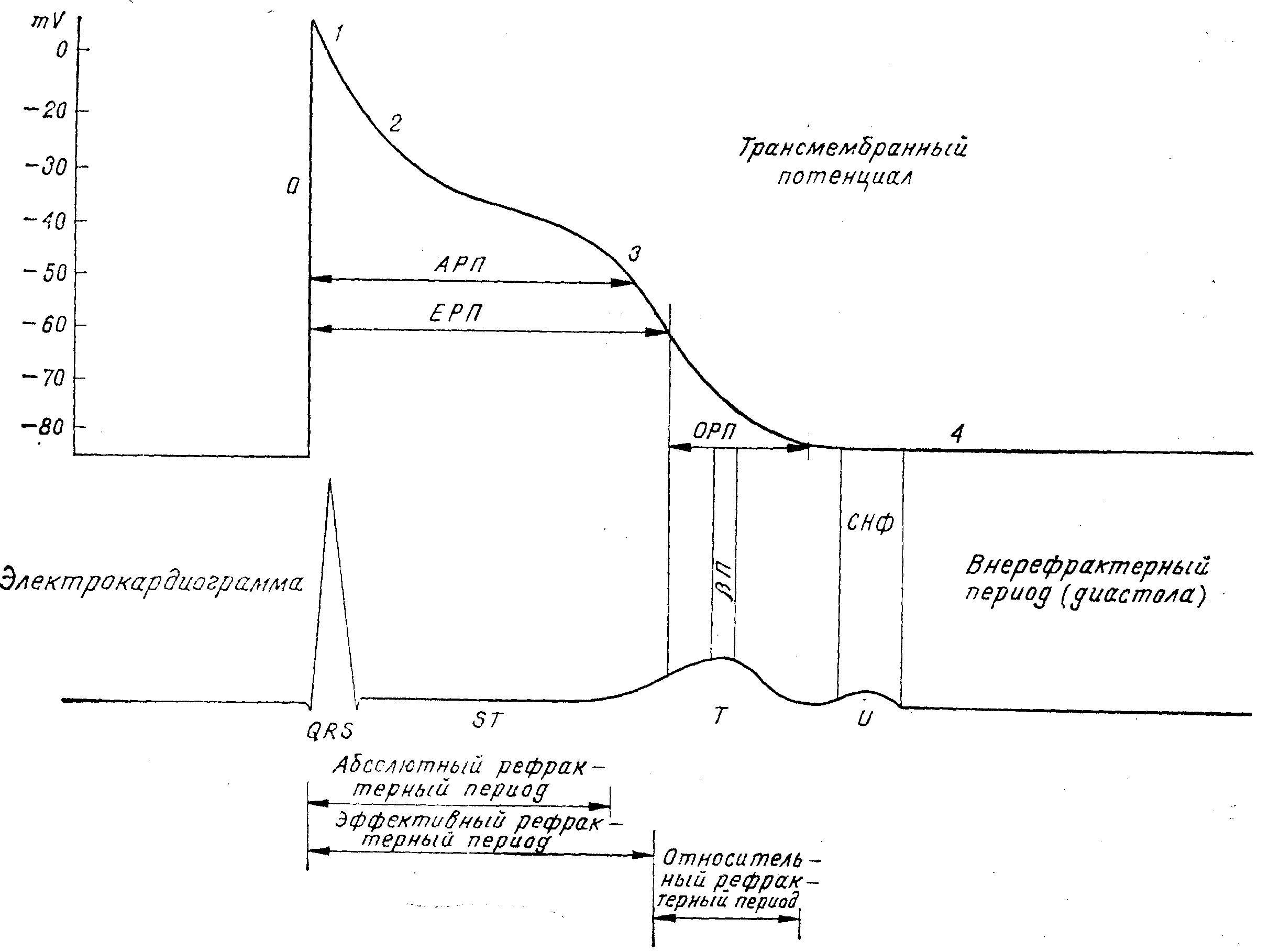
Рефрактерность сердечной мышцы.
АРП— абсолютный рефрактерный период; ЭРП — эффективный рефрактерный период; О^П—относительный рефрактерный период; ВП — вульнерабельный (уязвимый) период; СНФ — супернормальная фаза.
Во время абсолютного рефрактерного периода сердце не может активироваться и сокращаться, независимо от силы примененного раздражения.
Во время эффективного рефрактерного периода сердце способно активироваться, но полученный электрический импульс слабый и не распространяется, вследствие чего не наступает сокращения миокарда. Эффективный рефрактерный период охватывает абсолютный рефрактерный период и тот период, в течение которого возникает слабое электрическое активирование без распространения импульса. Вовремя относительного, релятивного или, называемого еще частичным, рефрактерного периода, сердце может активироваться при раздражении, более сильном, чем обычное. Полученный электрический импульс распространяется, хотя и медленнее чем нормально, и может привести к сокращению сердечной мышцы. Сумма эффективного и относительного рефрактерных периодов дает тотальный рефрактерный период. Тотальный рефрактерный период соответствует интервалу Q — Т на электрокардиограмме — электрической желудочковой систоле. Он соответствует всему потенциалу действия клетки. Абсолютный рефрактерный период соответствует комплексу QRS и начальной и средней части сегмента S—T на электрокардиограмме. Он охватывает потенциал действия с самого его начала до, примерно, —50 мв реполяризации. Конец абсолютного рефрактерного периода определяется как момент реполяризации, после чего при дополнительном раздражении может возникнуть слабый, нераспространяющнйся электрический импульс. Эффективный рефрактерный период соответствует комплексу QRS и всему сегменту S—T на электрокардиограмме. Он охватывает потенциал действия от его начала до, примерно, — 60 мв реполяризации. Конец эффективного рефрактерного периода определяется как момент реполяризации, вслед за которым при дополнительном раздражении может возникнуть медленно распространяющийся электрический импульс. Следовательно, разница между абсолютным и эффективным рефрактерным периодом заключается в том, что эффективный рефрактерный период охватывает также часть реполяризации, примерно, между—50 и—60 мв, когда при дополнительном раздражении может возникнуть слабый нераспространяющийся электрический импульс. Относительный рефрактерный период очень короткий и соответствует волне Т на электрокардиограмме. Он охватывает конечную часть реполяризации и находится приблизително между — 60 мв и концом потенциала действия.
Внерефрактерный период соответствует диастоле фазы 4 трансмембранного потенциала. В этот период проводниковая система и сердечная мышца восстанавливают возбудимость и способны к нормальному активнрованию.
Продолжительность рефрактерного периода различна в отдельных частях проводниковой системы и сократительного миокарда. Длиннее всего рефрактерный период в атриовентрикулярном узле. Среднее место по продолжительности рефрактерного периода занимает мышца желудочков, а предсердная мускулатура имеет самый короткий рефрактерный период. Правая ножка пучка Гиса имеет более длинный рефрактерный период, чем левая.
Продолжительность рефрактерного периода не постоянная величина. Она изменяется под влиянием многих факторов, но самое большое значение среди них имеет частота сердечной деятельности и вегетативная иннервация. Ускорение сердечной деятельности сокращает рефрактерный период, а замедление ее оказывает обратный эффект. Блуждающий нерв увеличивает продолжительность рефрактерного периода атриовентрикулярного узла, но укорачивает рефрактерный период предсердий. Симпатический нерв сокращает продолжительность рефрактерного периода всего сердца.
Существуют две, сравнительно короткие, фазы сердечного цикла, во время которых возбудимость сердца повышена: уязвимый (вульнерабельный) период и сверхнормальная фаза.
Уязвимый период находится в конечной части реполяризации и представляет собой составную относительного рефрактерного периода. Во время уязвимого периода пороговый потенциал понижен, а возбудимость клетки повышена. Вследствие этого, под воздействием даже сравнительно слабых раздражителей могут возникнуть желудочковые тахиаритмии и их мерцание. Ионный механизм этого периода не выяснен. Этот период приблизительно совпадает с пиком волны Т на электрограмме и соответствует небольшой части фазы 3 клеточной реполяризации.
Сверхнормальная фаза следует непосредственно после окончания относительного рефрактерного периода, соотв. реполяризации. Она находится в начале диастолы и часто совпадает с волной U на электрокардиограмме. Возбудимость сердечной клетки в этой фазе повышена. Незначительной силы раздражители могут вызвать необычно сильное электрическое активирование и тахиаритмии. Этот период обнаруживают только при функциональной депрессии сердца.
ИННЕРВАЦИЯ СЕРДЦА
Сердце иннервируется вегетативной нервной системой, регулирующей зарождение возбуждения и проведение импульсов. Она состоит из симпатических и парасимпатических нервов.
Преганглионарные симпатические волокна отходят от верхних 5 грудных сегментов спинного мозга. Они имеют синапсы в верхнем, среднем и нижнем шейных ганглиях и в звездчатом ганглии. От них отходят постганглионарные волокна, образующие симпатические сердечные нервы. Веточки этих нервов идут к синусовому и атриовентикулярному узлам, проводниковой ткани мышц предсердий и желудочков и венечным артериям. Эффект симпатического нерва осуществляется посредством медиатора норадреаналина, образующегося в окончаниях симпатических волокон в миокарде. Симпатические волокна увеличивают частоту сердечных сокращений и поэтому их называют cardioaccelerator.
Парасимпатические волокна сердце получает из блуждающего нерва, ядра которого расположены в продолговатом мозгу. От шейной части ствола блуждающего нерва отходят 1—2 веточки, а от грудной части — 3—4 веточки. Преганглионарные волокна имеют свои синапсы во внутристеночных ганглиях, расположенных в сердце. Постганглионарные волокна идут к синусовому и атриовентрикулярному узлам, предсердной мускулатуре, вверхней части пучка Гиса и венечным артериям. Наличие парасимпатических волокон в мышце желудочков еще не доказано. Медиатором парасимпатических волокон является ацетилхолин. Блуждающий нерв является кардиоингибитором: он замедляет сердечный ритм, оказывая тормозящее воздействие на синусовый и атриовентрикулярный узлы.
Афферентные нервные импульсы от кровеносных сосудов, дуги аорты и каротидного синуса проводятся в сердечно-сосудистый регуляторный центр в продолговатом мозгу, а эфферентные — от того же центра посредством парасимпатических и симпатических нервных волокон в синусовый узел и остальную часть проводниковой системы и коронарные сосуды.
РЕГУЛЯЦИЯ СЕРДЕЧНОГО РИТМА
Электрофизиологические процессы зарождения и проведения импульсов возбуждения в проводниковую систему и миокард находятся под влиянием ряда регуляторных нейрогуморальных факторов. Несмотря на то, что формирование импульсов в синусовом узле является автоматическим процессом, он находится под регулирующим влиянием центральной и вегетативной нервной системы. Синусовый и атриовентрикулярный узлы находятся исключительно под влиянием блуждающего нерва и в меньшей степени — симпатического. Желудочки контролируются только симпатическим нервом.
Дата добавления: 2016-07-11; просмотров: 9629;
Поиск по сайту
Узнать еще
- I.5.2 ПОТЕНЦИАЛ ПОЛЯ
- II группа действий. ВЗАИМОДЕЙСТВИЯ
- II. ОБУЧЕНИЕ ПРОСТЫМ БОЕВЫМ ДЕЙСТВИЯМ
- II. Получение вращающегося магнитного поля и принцип действия АД.
- II. Принцип действия и режимы работы синхронной машины
- II. Электрическая схема и принцип действия.
- III. БОЕВЫЕ ДЕЙСТВИЯ
- III. ОБУЧЕНИЕ СЛОЖНЫМ ДЕЙСТВИЯМ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
