Другие пути окисления одно- и двухуглеродных
Субстратов
Цикл трикарбоновых кислот осуществляется в клетках бактерий, грибов, простейших, растений, животных и является одним из самых важных циклов аэробных организмов. Однако у некоторых бактерий и растений существуют другие пути окисления одно- и двухуглеродных молекул. Как правило, использование альтернативных путей бывает вызвано тем, что в качестве единственного источника углерода и энергии используется какой-либо неуглеводный субстрат. В таком случае клетки вынуждены искать возможность такого способа окисления субстрата, который приводит к образованию промежуточных продуктов, способных превращаться в необходимые клетке метаболиты.
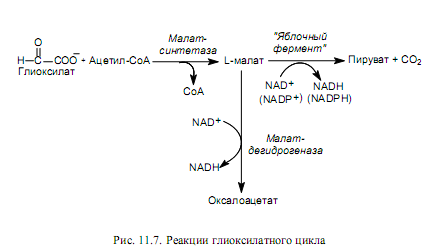
Примером подобной ситуации может служить функционирование глиоксилатного цикла в клетках семян высших растений. Когда основным запасным веществом семян служат триацилглицериды, остатки жирных кислот которых превращаются при окислении в ацетил-СоА, клетки используют глиоксилатный цикл (рис. 11.6, 11.7). Особенность этого цикла состоит в том, что изоцитрат не декарбоксилируется, как в ЦТК, а расщепляется под действием изоцитрат-лиазы на сукцинат (возвращается на этап b-окисления) и глиоксилат (рис. 11.6).
Глиоксилат затем конденсируется с еще одной молекулой ацетил-СоА (фермент малат-синтетаза) и превращается в L-малат. Это четырехуглеродное соединение может окисляться в оксалоацетат либо декарбоксилироваться с образованием пирувата. И оксалоацетат, и пируват могут использоваться клеткой в качестве предшественников биосинтеза многих нужных клетке веществ, в том числе глюкозы (рис. 11.7).
Этот процесс используют и бактерии, например E. coli, Pseudomonas, а также некоторые простейшие в случаях, когда единственным источни-
ком углерода служит ацетат. У бактерий ЦТК и глиоксилатный цикл пространственно на разделены, а у эукариот часть ферментов, необходимых для глиоксилатного пути, находится в особых органеллах —глиоксисомах. Глиоксисомы по структуре напоминают пероксисомы. Поскольку глиоксилатный путь можно рассматривать как анаплеротические реакции (восполняют количество сукцината, малата, оксалоацетата), то его часто считают частью ЦТК (рис.11.6).
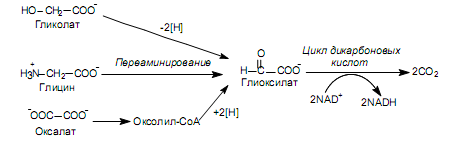
Некоторые бактерии существуют в условиях, когда единственным источником углерода являются такие двухуглеродные соединения, как гликолат, глицин, оксалат. Все эти вещества могут превращаться в глиоксилат (рис. 11.8), который, в свою очередь, окисляется до СО2 и Н2О в цикле дикарбоновых кислот. В этом цикле акцептором глиоксилата служит ацетил-СоА, который регенерируется в системе реакций. Каждый оборот цикла сопровождается образованием двух молекул углекислоты и высвобождением двух пар электронов, которые в составе двух молекул NADH поступают в дыхательную цепь.
Клетки многих микроорганизмов, а также некоторых растений и животных способны окислять и другие необычные субстраты, в том числе одноуглеродные (например, формиат), извлекая при этом энергию. В каждом случае реализуется оригинальный метаболический путь, позволяющий использовать отщепленные от субстрата электроны в дыхательной цепи для синтеза АТР.
Глава 12. ДЫХАНИЕ
Дыхание является одним из самых распространенных способов запасания энергии, которым обладает большинство организмов: животные, растения, простейшие, грибы, аэробные и почти все факультативно анаэробные бактерии. В процессе дыхания энергия запасается по механизму окислительного фосфорилирования, донорами электронов могут выступать как органические, так и неорганические вещества, а акцепторами электронов служат только неорганические вещества. По сравнению с субстратным окислительное фосфорилирование — гораздо более выгодный и эффективный механизм, в эволюционном плане находящийся на более высокой ступени. Его отличительной особенностью является обязательное участие мембран, в которых в строго определенной очередности расположены компоненты дыхательной цепи. Движущей силой синтеза АТР служит энергия протонного градиента на мембране. В свою очередь, протонный градиент создается в результате переноса электронов по компонентам дыхательной цепи в определенном направлении: от лучшего донора к лучшему акцептору.
Компоненты дыхательной цепи (электронтранспортная система) и катализирующий образование АТР фермент (АТР-синтаза) располагаются у прокариот в плазматической мембране, а у эукариот — во внутренней мембране митохондрий. Именно в этих органеллах, как было показано ранее (глава 9, 11), в ходе катаболических и амфиболических процессов формируется наибольшее количество восстановительных эквивалентов, которые с помощью никотинамидных и флавиновых переносчиков передаются в дыхательную цепь.
В дыхательной цепи осуществляются реакции, представляющие собой биохимический аналог горения водорода. Их отличительной особенностью является запасание значительной части выделяющейся энергии в виде макроэргических связей АТР, т. е. перевод свободной энергии в биологически доступную форму. И лишь небольшая доля выделяющейся при дыхании энергии рассеивается в виде тепла.
Для понимания механизма окислительного фосфорилирования необходимо, прежде всего, охарактеризовать компоненты дыхательной цепи, закономерности их функционирования и расположения в мембране.
Дата добавления: 2016-05-30; просмотров: 2278;
Поиск по сайту
Узнать еще
- F38 Другие (аффективные) расстройства настроения
- F68 Другие расстройства зрелой личности и поведения у взрослых
- II. Судовождение с использованием лоцманского метода и графического счисления пути судна.
- III и IV нейроны слухового пути. Третьи и четвертые нейроны слухового проводящего пути. Ядра слухового анализатора. Признаки поражения слухового пути.
- III. Реакции окисления
- IV. Ликвидация гиповолемии и регуляция водно-солевого обмена
- IV. проводящие пути головного и спинного мозга
- Pиc. 67. Зависимость скорости осаждения от величины катодно-анодного отношения

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
