Предмет экологии насекомых 11 глава
Следовательно, как правило, одна популяция живет в пределах одного биотопа. Если же граница этого биотопа не может быть определена четко, мы не можем отделить одну популяцию от другой, как навряд ли возможно разграничить отдельные популяции насекомых, обитающих в тайге или нетронутой степи.
Тем не менее даже при четком разграничении биотопов, но сравнило небольшом расстоянии между ними возможен интенсивный обмен мигрирующими особями. Поэтому все близко расположенные друг к другу популяции составляют континуум, распространяющийся на большой географический район с более или менее однородными климатическими условиями и сообществами растений. Такую совокупность элементарных популяций называют суперпопуляцией. Тогда сплошной ареал вида может быть представлен как континуум соприкасающихся суперпопуляций, а разорванный ареал – как совокупность изолированных суперпопуляций.
Иногда предполагается, что элементарная популяция в границах биотопа тоже подразделена на так называемые микропопуляции, т.е. населения одного или нескольких находящихся рядом деревьев или даже одного пня. Такое разграничение обычно не оправдано и возможно только при малой подвижности насекомых. Если же насекомые интенсивно передвигаются в пределах биотопа, то различия в генофонде населения отдельных микроучастков отсутствуют или случайны. Однако, благодаря различиям по микроклимату или кормовым свойствам отдельных растений, насекомые, развивающиеся на таких участках, могут заметно отличаться друг от друга по размерам, морфологии или окраске.
Итак, в пределах вида возникают следующие совокупности особей, все более тесно связанных общей генетической информацией друг с другом: вид (ареал) – суперпопуляция (географический район) – элементарная популяция (биотоп) – микропопуляция (микробиотоп).
Географическая популяция – аллопатрическая дивергенция
Очевидно, что, с одной стороны, совокупности особей в разных популяциях могут отличаться друг от друга по случайным причинам в зависимости от генофонда особей, основавших эти популяции. Возможность таких различии резко увеличивается при большом удалении друг от друга биотопов, пригодных для жизни насекомых этого вида, или наличии трудно преодолимых преград, например горных хребтов или на островах. Если же преграды для расселения отсутствуют и недалеко находятся другие популяции этого же вида, постоянный приток мигрантов будет нивелировать случайные отклонения, связанные с генофондом основателей, и отличия одной популяции от другой будут продиктованы прежде всего особенностями условий в том или ином биотопе.
С другой стороны, чем дальше друг от друга расположены биотопы, тем больше различаются их климат, почвы и растительные ассоциации. Соответственно, благодаря естественному отбору между отдаленными друг от друга популяциями могут возникнуть существенные различия. При сравнении групп насекомых, взятых из географических пунктов, отдаленных друг от друга на сотни километров, принято говорить о географических популяциях.
Итак, благодаря "эффекту основателя", возникающему при отсутствии обмена генетической информацией, отбору применительно к местным условиям, а также случайному возникновению адаптивных мутаций в удаленных друг от друга популяциях, неизбежно возникают более или менее заметные различия в биологии, морфологии и поведении (аллопатрическая дивергенция). Эта дивергенция может привести к тому, что обмен генетической информацией окажется невозможным даже при случайном залете мигрантов из-за морфологического несоответствия особей, расхождений в их брачном поведении или же нежизнеспособности потомства.
Различия между географическими популяциями возможны по любым признакам, в том числе по фотопериодической реакции (критический период реакции щавелевой совки закономерно увеличивается в ряду популяций Сухуми–Белгород–Витебск–Ленинград) и по термопреферендуму (даже у такого синантропного вида, как комнатная муха, есть определенные различия в термопреферендуме северных и южных популяций).
Очень интересны особенности географического полиморфизма у бабочки Papilio dardanus, распространенной почти по всему африканскому континенту. Эти бабочки подражают внешнему облику и окраске других местных бабочек, несъедобных для животных. В каждом географическом районе обнаруживается до 7–10 моделей для подражания. Такая мимикрия приводит к громадному разнообразию этого вида при резко очерченных внутривидовых формах.
Известны и такие относительно редкие случаи, когда особи, живущие в пределах одного биотопа, отличаются друг от друга в такой же степени, как представители разных географических популяций. Именно такая картина изменчивости была выявлена у некоторых наездников–яйцеедов (Пала Рам, 1990). Подобное отклонение от общего правила, по–видимому, связано с тем, что у этих видов резко нарушена панмиксия, т.е. возможность свободного скрещивания между особями. Действительно, у многих перепончатокрылых паразитов, в том числе у трихограммы, одну кладку яиц хозяина заражает, как правило, одна самка. Выведшиеся из кладки сестры и братья (сибсы) спариваются друг с другом. Вероятность встречи брачных партнеров из разных кладок хозяина очень мала. При таком инбридинге популяции складываются из отдельных более или менее чистых симпатрических природных линий, как при партеногенезе или вегетативном размножении.
Экологические расы – парапатрическая дивергенция
Две различные популяции могут существовать и бок о бок в том случае, если они приспособлены к двум заметно отличающимся друг от друга биотопам. Так, на песчаных и лессовых почвах обитают несколько различные жуки–чернотелки, относящиеся к одному виду – Tentyria nomas Pall. В лаборатории эти жуки в 2/3 случаев предпочитали спариваться с особями "своей" популяции (К.В.Арнольди, 1941).
Удивительно разделение двух рас мух Drosophila funebris F.: городской и деревенской. У дрозофил этого вида, отловленных в разных частях Москвы, частота гетерозигот по одной инверсии равна 50–88%, а в 10–12 км от Москвы – всего 1–2%. Между этими зонами имеет место резкий перепад частоты инверсий.
Такие популяции одного вида, занимающие соседние территории с разными типами местообитаний и сохраняющие свою самостоятельность без наличия каких–либо внешних препятствий, называют экологическими расами.
Сезонные расы – симпатрическая дивергенция
В пределах одного биотопа может сосуществовать несколько сезонных рас, мало или совсем не обменивающихся генетической информацией. Так, у сверчка Gryllus firmus наиболее обычна зимовка на стадии яйца и появление половозрелых особей к началу осени. Однако некоторые сверчки того же вида в том же биотопе зимуют на стадии личинки и превращаются в имаго уже весной. Никакой обмен генетической информацией между этими расами невозможен. Аналогичным образом могут формироваться и сезонные расы у ряда саранчовых.
Возможны и не столь резкие различия в сезонности, тем не менее приводящие к довольно полной генетической изоляции. Так, одуванчиковые долгоносики Ceutorrhynchus optator на склоне оврага с северной экспозицией появляются на 15–20 дней позже, чем на противоположной стороне. Иногда различия во времени появления половозрелых особей связаны с использованием различных кормовых растений (плодовые мухи Rhagoletis).
Биологические расы – симпатрическая дивергенция
Помимо изменений сроков развития, питание фитофагов на различных кормовых растениях может приводить и к другим затруднениям в передаче генетической информации. Согласно правилу Гопкинса (A.D.Hopkins, 1917), многие насекомые фитофаги предпочитают откладывать яйца на растения того же вида, которым они питались на стадии личинки. По–видимому, здесь имеет место запоминание насекомым запаха или других свойств кормового растения, аналогичное запечатлению (импринтингу). Подобное же явление возможно при развитии одного и того же паразита на разных хозяевах. Таким образом, ряд насекомых способны сохранять память о корме на протяжении метаморфоза, хотя у преобладающего большинства видов обучение происходит только на какой-то определенной стадии (С. Я. Резник, 1993). Если при этом спаривание насекомых преимущественно происходит на хозяине или около него или же возникают изменения в окраске и поведении, связанные с различным кормом, то в пределах популяции дифференцируются биологические расы. Как правило, эти расы генетически не полностью изолированы друг от друга.
Биологические расы подробно изучены у ивового листоеда Lochmaea caprea L. Известны две расы этих жуков, обитающие в одних и тех же биотопах: ивовая, питающаяся на разных видах ив, и березовая, потребляющая листья пушистой березы. Внешне жуки этих двух рас практически неразличимы. Отмечены лишь определенные различия в их отношении к микроклимату (А.В.Михеев, А.Г.Креславский, 1980). Однако жуки ивовой расы не способны или почти не способны питаться на березе, в то время как "березовые" жуки одинаково охотно питаются и березой, и ивой. Правда, в последнем случае у личинок повышается смертность. При скрещивании жуков этих двух рас потомство способно с одинаковым успехом питаться и березой, и ивой. Расчет, однако, показывает, что такие скрещивания в природе редки – не более 5% случаев (А.Г.Креславский и др., 1981). Интересно, что у другого вида листоедов – Chrysochloa calcaliae Schrnk., хотя и намечается предпочтение определенными жуками определенных растений, дифференциация гораздо слабее (В.М.Соломатин и др., 1976). В какой-то мере это объясняется тем, что спаривание у Chrysochloa имеет место не на кормовых растениях, как у ивового листоеда, а на проталинах, где собираются все жуки, независимо от кормового растения, на котором они питались.
Специализированные по растениям биологические расы известны и у ряда других насекомых. Особенно четко они выражены у тлей, где каждая биологическая раса характеризуется своим набором расцветок. Тли с трудом переходят на новое для них растение, однако между этими расами могут быть образованы гибриды.
Вопрос о специализации паразитов по отношению к их хозяевам имеет большое значение для практики биометода. Если такая специализация имеет место, необходимо разводить для выпуска на поля не любых, случайно паразитов, а совершенно определенную биологическую расу, приспособлению к данному хозяину. Так, наездники–яйцееды каждого вида Trichogramma способны заражать яйца многих хозяев, преимущественно бабочек. Кладки бабочек обычно не встречаются в массе, и в природе их трудно найти, тем более, что исключительно малые размеры этих наездников (длина тела около 0,5–0,7 мм), а также относительно слабо развитые органы чувств затрудняют поиск хозяина. Кроме того, цикл развития трихограммы гораздо короче, чем циклы развития бабочек – ее хозяев. Поэтому чрезмерно узкая специализация трихограммы по отношению к определенным видам хозяев привела бы этого паразита к вымиранию. И тем не менее у трихограмм одного и того же вида обнаруживается врожденное и различное предпочтение определенных хозяев (Пала Рам,1990). В любом случае, однако, крупные яйца охотнее заражаются, чем мелкие а наиболее привлекательными для большинства видов трихограмм оказываются яйца бабочек–совок (А.П.Сорокина, 1987).
Полиморфизм в популяциях
Неоднородность генотипа и фенотипа особей, входящих в популяцию, имеет большое экологическое значение. В зависимости от конкретных условий больше шансов выжить то у одних, то у других особей, которые затем снова восстанавливают всю популяцию. Так, например колорадский жук способен быстро наращивать свою численность даже после воздействия очень неблагоприятных условий: сплошных химических обработок или временного отсутствия своего кормового растения, благодаря разной продолжительности диапаузы у разных особей. Диапаузирующие жуки могут находиться в почве от одного года до трех лет.
Иногда эта изменчивость может регулироваться внешними факторами, как бы направляющими особь по тому или иному пути развития. Так под влиянием большой плотности популяции саранча Locusta migratoria L. существенно меняет свой облик и физиологию, переходя из одиночной фазы в мигрирующую. Мигрирующая саранча охотно образует скопления – кулиги личинок и стаи имаго, передвигающиеся в поисках корма. Плодовитость мигрирующей саранчи в 3–4 раза ниже, чем одиночной.
Некоторую аналогию с мигрирующей саранчой представляют крылатые особи тлей, возникающие, в частности, при высокой скученности тлей на растении или ухудшении питания. Правда, крылатые тли, не способные к активному полету из-за своих малых размеров, мигрируют одиночно, не образуя скоплений. Под влиянием условий меняется соотношение длинно- и короткокрылых особей одного вида клопов–водомерок. Жаркое и сухое лето способствует появлению большого количества длиннокрылых мигрирующих особей, что необходимо в связи с массовым пересыханием мелких водоемов. Короткокрылые же особи оказываются более устойчивыми к неблагоприятным условиям и могут вытеснять длиннокрылых.
Полиморфизм может быть обусловлен и чисто генетическими причинами. Так, у двуточечной божьей коровки Adalia bipunctata L. известны две формы: черная и светло–оранжевая. Черные жуки, по крайней мере, в некоторых географических районах, отличаются большей смертностью при зимовке, что, по-видимому, связано с повышенной отдачей тепла при такой окраске. Эта смертность компенсируется большей половой активностью черных особей, благодаря чему в природных популяциях поддерживается относительно стабильное равновесие.
В принципе, меланистические темные формы получают адаптивно преимущество, если темнеет "окружающий фон". Так, в индустриальных районах Англии заметно возрастает количество окрашенных в черный цвет или хотя бы более темных березовых пядениц Biston betularia L., что связывается с более темными и лишенными лишайников стволами деревьев при существенном загрязнении воздуха.
Изменчивость в популяции может быть и не так очевидна, но имеет большое приспособительное значение. В любой популяции можно найти более и менее склонных к миграции особей. В зависимости от запасов пищи и погоды преимущественно выживает та или иная часть популяции, сохраняя ее общий генофонд. Кроме того, в популяции всегда выделяются менее разборчивые к питанию или другим условиям особи и особи, более специализированные по своим потребностям и возможностям (А.Г.Креславский, 1987).
Итак, возможность выживания популяции резко увеличивается благодаря ее генетической и фенетической неоднородности
Основание культуры насекомых
Очевидно, что каждая культура насекомых в лаборатории или на биофабрике, полностью соответствует определению популяции. При основании культуры насекомых, как правило, мы имеем дело с относительно небольшим количеством насекомых-основателей. Поэтому исключительно важно выбрать наиболее подходящую для поставленной задачи географическую популяцию, а в пределах этой популяции найти биологическую расу с необходимой специализацией. Вероятность удачного выбора исходного материала значительно повышается в начале вспышек численности насекомых, если таковые имеют место.
Ограниченность исходной выборки не позволяет исчерпать весь генофонд популяции, поэтому желательно иметь, возможно, больше отдельных экспериментальных "линий", чтобы выбрать из них лучшие. Обычно такие линии представляют собой потомство наиболее перспективных по своим признакам самок с введением на некоторое время инбридинга. Отметим, что инбридинг далеко не всегда отрицательно влияет на свойства культуры.
Изменения генофонда популяций
Согласно известному правилу Харди–Вайнберга, частоты генов в популяции в течение определенного времени достигают равновесия и далее их соотношение в генофонде популяции остается неизменным. Однако это возможно лишь при условии, что вообще отсутствует какой–либо отбор или миграции особей с определенными свойствами, а скрещивание между особями происходит случайно (абсолютная панмиксия). Кроме того, численность такой популяции должна быть бесконечно большой. Очевидно, что в природе эти условия выполнимы лишь в определенной степени, а это означает, что генофонд природной популяции не может быть абсолютно стабильным.
Генофонд может обогащаться за счет прибытия мигрантов из других популяций, а также за счет мутаций. Концентрации тех или иных аллелей могут меняться в результате воздействия внешних условий, приводящих к гибели или снижению плодовитости особей с тем или иным генотипом, т.е. в результате отбора. Правда, при прекращении отбора в случае сохранения гетерозиготности восстанавливаются прежние частоты генов. Генофонд, популяции может быть обеднен при регулярной утечке обладающих определенными свойствами мигрантов.
Другая причина обеднения генофонда связана с так называемым дрейфом генов при малой численности популяции. Механизм этого явления достаточно четко описан в "Курсе общей энтомологии" Ю. А. Захватами (1986). Приведем здесь отрывок из этой книги.
"В основу концепции дрейфа генов положены представления о случайном характере распределения частот генов и непредставительности (не презентативности) малых выборок. Обратимся к обычному для статистиков примеру. Из урны, заполненной 5000 белыми и 5000 черными шарам (символизируют для нас аллели А и а), вынем подряд первую тысячу шаров и убедимся, что около половины из них будут белыми. Допустим, что мы вынули 514 белых и 486 черных шаров. Увеличив каждое из этих чисел в 10 раз (размножение – В.Ч.), заполним вторую урну 5140 белыми и 4860 черными шарами и, перемешав их, повторим процедуру. Возможно, что из 1000 взятых на этот раз шаров 506 окажутся черными, а 494 белыми. Многократно повторив данный эксперимент, можно убедиться, что число шаров разного цвета каждый раз будет близким к 500.
Однако если мы заполним урну только 100 шарами и вынем первые 10, то, возможно, среди них белых (или черных) будет несколько больше половины, например 6. Теперь возьмем 60 белых и 40 черных шаров и, перемешав их в урне, вынем снова первые 10. Скорее всего, число белы шаров будет большим и, многократно повторяя эту процедуру, мы увидим что число черных шаров становится все меньшим и, наконец, они все будут замещены только белыми. Неизбежное следствие наблюдаемого при этом "дрейфа генов" – утрата одних и фиксация других аллелей, а это, в свою очередь, приводит к сокращению гетерозиготности популяций и затуханию связанной с этим изменчивости. Скорость этого затухания прямо пропорциональна величине популяции, чем она меньше, тем быстрее проявляются все последствия дрейфа".
Действительно, практика разведения насекомых показывает, что дрейф генов становится заметным лишь при исходной численности популяции менее 100 особей. Если же исходная численность более 500, дрейфом генов можно пренебречь.
Очевидно, что при культивировании насекомых во многих случая можно обнаружить дрейф генов и соответствующее уменьшение гетерозиготности. Сохранять гетерозиготность по генам, не имеющим существенного значения для необходимых свойств культуры, и тем самым предотвращать ее возможное вырождение можно, сочетая инбридинг с аутбридингом (неродственным спариванием). Однако это целесообразно лишь в том случае, если гетерозис связан со сверхдоминированием по генам, влияющих на приспособленность. Если же инбредная депрессия обусловлена тем, что вредные рецессивные мутации становятся гомозиготными, максимальный эффект достигается отбором наилучших инбредных линий. Во многих же случаях дрейф генов вообще не проявляется, так как ему противостоит отбор по адаптивным признакам.
Подчеркнем, что дрейф генов может иметь место и в природе при существенном сокращении численности популяций по любым причинам: от неблагоприятной погоды, эпизоотии, применения инсектицидов, или любых других защитных мероприятий. Возрождающаяся после таких воздействий популяция имеет уже несколько иной генофонд, причем изменения популяции направляются отбором в сторону наибольшей устойчивости к этим неблагоприятным воздействиям. Аналогичная потеря богатства генофонда, вызванная дрейфом генов, имеет место в природе при заселении новой территории незначительным количеством мигрантов. Однако если поток мигрантов продолжается, происходит встречное обогащение генофонда
Другая причина вырождения лабораторной культуры – это лабораторные условия, как правило, обеспечивающие сохранение и размножение ослабленных особей с неблагоприятным генотипом, которые неминуемо погибли бы в природе. Более того, эти ослабленные особи получают преимущество в размножении, так как они обычно менее агрессивны и лучше переносят высокую плотность популяции, которая в лаборатории обычно на несколько порядков выше, чем в природе,
Тем не менее известно много случаев, когда лабораторная культура стабилизируется и может существовать на протяжении неопределенного числа поколений. Как мы отмечали выше, здесь многое зависит от того, насколько удачно взят исходный материал. Во многих же случаях после 3–4 поколений культура насекомых погибает и приходится брать новый материал.
Популяции насекомых при смене корма
В природе иногда возникает необходимость замены одного корма другим. Так, случайно упавшие с ольхи на землю личинки листоеда Chrysomela aenea L. не всегда могут вернуться назад в крону и часто заканчивают свое развитие, питаясь крапивой. Однако взрослые жуки, по–видимому, никогда не откладывают яйца на крапиву.
Подобная задача возникает при попадании насекомого в новый для него географический район с иной флорой. Иногда при введении новых сельскохозяйственных растений местные насекомые осваивают необычный для них корм, который в ряде случаев оказывается более благоприятным чем исходный (например, переход колорадского жука на культурные сорта картофеля, клопа–черепашки на полиплоидные сорта пшеницы).
В опытах Г.Х.Шапошникова (1966) тли при переходе на новые и исходно малопригодные для них растения за 8 поколений полностью адаптировались к ним. Эти тли оказывались неспособными скрещиваться с исходной контрольной линией, а также питаться на исходном растении. Подобный процесс адаптации тлей к новому растению подробно описан Е.С.Смирновым (1961), который наблюдал "волнообразные" изменения признаков от поколения к поколению, затухающие после полной адаптации и выхода лабораторной популяции на новый уровень.
Чаще всего, однако, при питании новым кормом снижаются жизнедеятельность и плодовитость насекомых, происходит уменьшение их размеров. В частности, такая картина проявляется во многих случаях при использовании искусственных питательных сред (А.З.Злотин, 1989).
Трихограмма в природе преимущественно заражает яйца бабочек совок. В ряде стран (Россия, Украина, США) в качестве лабораторного хозяина используют зерновую моль – вид, с которым трихограмма никого не встречается в природе. В принципе, трихограмма может развиваться на этом хозяине сколь угодное число поколений. Правда, начиная с первого поколения на ситотроге, размеры трихограммы уменьшаются (по–видимому, из-за малых размеров этого хозяина), но дальше остаются стабильными.
Однако, такая трихограмма пригодна для выпуска в поле только не более чем на протяжении первых трех поколений, далее же эффективность этого энтомофага резко снижается. Наблюдения показывают, что с первого и до четвертого поколения происходит резкое снижение способности трихограммы к прыжкам и полету. Начиная с пятого поколения на ситотроге эти способы передвижения можно наблюдать только при очень высокой освещенности и высокой температуре. Обратный перевод трихограммы с ситотроги на естественного хозяина – капустную совку в первом же поколении восстанавливает способность передвигаться прыжками. При выращивании на ситотроге также пропадает способность самок реагировать на кайромоны – вещества, сохраняющие запах естественного хозяина на его кладке яиц и поблизости от нее, которые помогают наезднику обнаружить эту кладку (Л.М.Салманова и др.,1992).
Скорее всего, такие изменения связаны не с отбором особей, наиболее приспособленных к новому хозяину, а с так называемыми длительными модификациями. Механизм этого явления здесь может заключаться в постепенном обеднении питательными веществами яиц, откладываемых самками яйцееда последовательных поколений.
2. ХАРАКТЕР РАЗМЕЩЕНИЯ НАСЕКОМЫХ НА МЕСТНОСТИ
Размещение особей одного вида может быть трех типов: равномерное, случайное и агрегированное (рис.20). Эти типы размещения плавно переходят из одного в другой и четкой границы между ними нет. Очевидно, что, с одной стороны, размещение может определяться степенью однородности заселяемого участка по микроклимату и растительности, с другой – быть связано с ограниченностью пищевых ресурсов, а также взаимодействиями между особями. Характеристика размещения популяции на местности – важнейшая сторона экологии и поведения изучаемого вида. Кроме того, без знания типа размещения особей невозможен точный учет численности насекомого – характеристики популяции, имеющей большое практическое значение.
При изучении размещения насекомых на местности следует учитывать, что характер размещения насекомых зависит не только от вида, но и от плотности его популяции. Например, малые мучные хрущаки Tribolium confusum Duv. при малой плотности образуют скопления в субстрате (агрегированное размещение), с увеличением плотности они размещаются в субстрате случайно и, наконец, при высокой плотности их размещение становится равномерным.
Как правило, размещение насекомых на местности не может быть определено визуально. Для этого берут почвенные пробы, производят учет на отдельных растениях или отдельных ветках деревьев. Однако результат определения типа размещения может зависеть от размера пробы. При неудачно выбранных слишком больших или слишком маленьких размерах проб агрегированное размещение может быть принято за случайное.
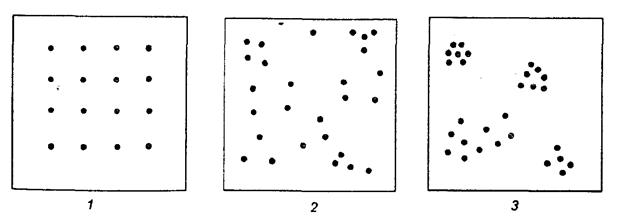
Рис.20. Основные типы размещения особей: I – равномерное или регулярное, 2 – случайное, 3 – агрегированное или пятнистое (по А.В.Смурову, 1975)
Ориентировочно характер размещения может быть определен по коэффициенту вариации числа особей в пробах δ 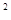 /
/ 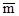 , где δ – дисперсия, а – арифметическое среднее. Если этот коэффициент меньше единицы, размещение может считаться равномерным, примерно равен единице (0,8– 1,2) – случайным, если больше – размещение агрегированное. Более точный результат может дать сравнение теоретического ряда, соответствующего определенному типу статистического распределения, и эмпирического, полученного при подсчете особей в пробах с помощью статистического критерия χ . При этом желательно, чтобы подобранное таким образом теоретическое статистическое распределение имело определенный биологический смысл.
, где δ – дисперсия, а – арифметическое среднее. Если этот коэффициент меньше единицы, размещение может считаться равномерным, примерно равен единице (0,8– 1,2) – случайным, если больше – размещение агрегированное. Более точный результат может дать сравнение теоретического ряда, соответствующего определенному типу статистического распределения, и эмпирического, полученного при подсчете особей в пробах с помощью статистического критерия χ . При этом желательно, чтобы подобранное таким образом теоретическое статистическое распределение имело определенный биологический смысл.
Равномерное размещение
Такое размещение встречается редко, так как возможно лишь в относительно однородной среде. Оно обычно вызвано резко конкурентными отношениями между особями, их взаимным "отталкиванием" и наиболее вероятно у насекомых с четко выраженной К-стратегией, т.е. таких насекомых, которые жестко привязаны к определенному биоценозу и возможности их размножения ограничены. Так, относительно равномерно распределены некоторые хищные насекомые, активно оберегающие свою территорию (стрекозы, ктыри). Равномерное размещение может проявиться также и при очень большой плотности населения, когда отсутствует или резко подавлена возможность эмигрировать, например, в лабораторных культурах. Однако такое размещение возникает только в том случае, если механическое раздражение, запах или вид другой особи отталкивают их друг от друга. Как мы отмечали выше, именно так распределены в субстрате личинки малого мучного хрущака при большой плотности популяции.
При равномерном размещении каждая особь занимает примерно одинаковую площадь. Тогда, если размеры пробы значительно больше, чем эта площадь, количество особей в пробах будет примерно постоянным и близким к арифметической средней:
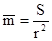
где S-площадь пробы, а r-среднее расстояние между особями. Итак, в случае равномерного размещения дисперсия между пробами относительно мала и, соответственно, отношение 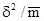 всегда меньше единицы.
всегда меньше единицы.
При размере же проб меньше средней площади, занимаемой особью, анализ покажет случайное распределение, соответствующее формуле Пуассона, которое описано в следующем разделе.
Равномерное размещение на местности хорошо описывается теоретическим биномиальным распределение (А.В.Смуров, Л.В.Полищук, 1989). Биологический смысл биномиального распределения здесь будет заключаться в следующем.
Предположим, что минимальная территория, занимаемая одной особью, равна s. Естественно, что по тем или иным причинам между особями, скорее всего, будут незанятые места – "пустые" пространства. Представим себе, что каждая проба захватывает площадь S, равную s, и границы каждой пробы точно совпадают с границами территорий, принадлежащих особям. Тогда часть проб будет содержать по одной особи, а часть проб будет пустыми. Например, если занята половина территории, то наиболее вероятно, что в половине наших проб будет по одной особи, а в половине проб не будет ни одной особи. В общем же случае, если вероятность того, что территория занята, равна p, а не занята – q, где p + q = 1, то отношение числа проб с одной особью к числу пустых равно p : q.
Дата добавления: 2016-06-29; просмотров: 1700;
Поиск по сайту
Узнать еще
- I.1.1 ПРЕДМЕТ МОЛЕКУЛЯРНОЙ ФИЗИКИ
- Активные – воздействуют на предмет труда, перемещают его, контролируют ход производства (машины, оборудование, т/с, инструмент)
- Актуальность изучения экологии
- Алфавитно-предметный указатель
- Аналитическая химия и химический анализ. Предмет и задачи аналитической химии. Классификация методов химического анализа.
- Аналогично работают ПТК СВШД и системы обнаружения случайных предметов (СОСП).
- Анатомо-физиологические аспекты потребностей человека. Задачи изучаемого предмета.
- Антропогенное влияние на природу. Проблема экологии.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
