Явления компромисса (баланса) в поведении животных
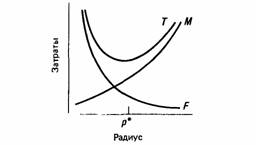
Большинство конструкторских решений требует компромисса между теми преимуществами, которые можно извлечь из несовместимых друг с другом особенностей данной ситуации. Так, например, если рассмотреть конструкцию простого кровеносного сосуда, то окажется, что затраты энергии на трение при протекании крови через сосуд уменьшаются с увеличением радиуса сосуда, тогда как затраты энергии на поддержание тонуса сосудистой стенки возрастают (рис. 24.5). Такой тип прямого сравнения затрат и выгод называется компромиссом (tradeoff). Удачной обычно бывает такая конструкция, в которой используются балансовые зависимости, присущие данной ситуации. Отсюда следует, что выявление балансовых зависимостей в поведении животных может служить хорошей отправной точкой при анализе того, таким образом действует естественный отбор.
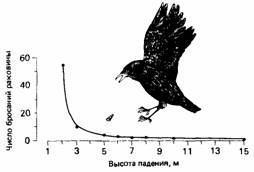
Зах (Zach, 1979) выявил простой пример баланса в пищедобывательном поведении ворон, которые питаются брюхоногими моллюсками на западном побережье Канады. Во время отлива они ловят моллюсков, выбирая обычно самых крупных из них. Отыскав такого моллюска, ворона поднимается с ним в воздух над скалистым берегом, а затем бросает раковину вниз; раковина разбивается, и ее съедобное содержимое становится доступно птице. Бросая моллюсков различных размеров с разной высоты, Зах обнаружил, что число бросаний, необходимое для того, чтобы раковина разбилась, зависит от высоты, с которой бросают раковину (рис. 24.6). Воронам приходится расходовать энергию для того, чтобы взлететь на какую-то высоту и бросить моллюска. Поэтому Зах рассчитал общее количество взлетов, которое должна была сделать ворона, чтобы разбить раковину, бросая ее с данной высоты. Оказалось, что расход энергии на взлеты будет наименьшим, если раковину бросать с высоты около 5 м. Это хорошо видно на рис. 24.7. Как и в конструкции кровеносного сосуда, здесь мы встречаемся с балансом - на этот раз
| 
|
| Рис. 24.5. Энергетические затраты на поддержание тока крови в зависимости от радиуса кровеносного сосуда. F - расход энергии на трение; М расход энергии на поддержание тонуса стенки сосуда; Т-- общие энергозатраты; р* - оптимальный радиус сосуда, при котором величина Т минимальна (По Milsum, Roberge, 1973.) | Рис. 24.6. Число бросаний раковины, необходимых для того, чтобы она разбилась, в зависимости от высоты падения раковины. (По Zach, 1979.) |
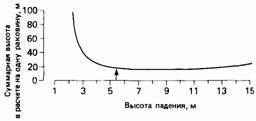
между затратами в виде числа бросаний раковины, требующихся для того, чтобы ее разбить, и высотой бросания. Расчеты, проведенные на основе этого баланса, показали, что существует некоторая оптимальная высота бросания (рис. 24.7), аналогичная оптимальному радиусу кровеносного сосуда (см. рис. 24.5). Зах обнаружил, что высота, с которой вороны обычно бросают раковины, близко соответствует расчетной оптимальной высоте бросания, равной 5 м. Таким образом, создается впечатление, что в поведении ворон заложена какая-то программа, которая использует характерные особенности той ситуации, в которой им приходится добывать пищу.
Расчеты затрат и выгод, которые относятся к питающимся моллюсками воронам, так же как и расчеты конструкции кровеносных сосудов, очень упрощает то обстоятельство, что все балансируемые факторы можно измерить с точки зрения затрат энергии. Расчеты Заха дают основание заключить, что вороны выбирают раковины определенного размера и бросают их с определенной высоты, получая при этом наибольший выигрыш в чистой энергии. Однако не всегда возможно или даже желательно рассматривать оптимальный баланс только с энергетической точки зрения. Как мы уже видели в случае травника, энергия не всегда играет определяющую роль. Более того, животные далеко не всегда имеют возможность сконцентрировать всю свою активность исключительно на добывании пищи. Им приходится еще и остерегаться хищников, и охранять территорию от соперников и т. д. Поэтому необходимо рассматривать ситуацию в целом, не забывая, что добывание пищи - это всего лишь один из видов активности в поведенческом репертуаре животного и что затраты или приобретение энергии не единственный результат поведения.
У большой синицы (Partis major) песня играет важную роль в защите территории; кроме того, она используется, чтобы заявить о себе (Krebs, 1971). Ранней весной птица поет в основном на рассвете, начиная свою песню в тот момент, когда солнце должно вот-вот показаться на горизонте. Такое хоровое пение ранним утром типично для многих птиц. Однако в этом есть что-то непонятное: ведь маленькие птички в большинстве своем теряют в течение морозной ночи до 10% массы тела, и поэтому, проснувшись, они должны испытывать сильный голод. Почему же тогда по утрам они прежде всего начинают петь, а не добывать пищу? По-видимому, для этого существуют две основные причины. Во-первых, защита территории особенно важна сразу после рассвета, поскольку именно в это время не имеющие территории птицы проверяют прочность «линии обороны», чтобы обнаружить какой-нибудь участок, освободившийся накануне в результате деятельности хищников.
|
| Рис. 24.7. Суммарная высота взлета вороны (произведение числа бросаний на высоту падения раковины) минимальна при такой высоте (отмечена стрелкой), с которой обычно вороны бросают раковины. (По Zach, 1979.) |
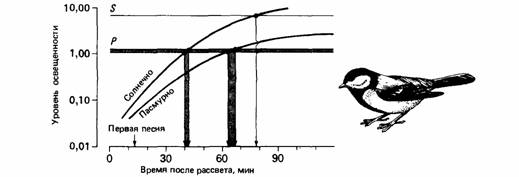
Во-вторых, выгоды от добывания пищи в это время невелики, потому что при слабой освещенности эта деятельность оказывается малоэффективной. Каселник (Kacelnik, 1979) в лабораторных экспериментах показал, что большие синицы не могут эффективно добывать пищу при освещенности ниже определенного уровня, а этот уровень достигается обычно через 40-80 мин после рассвета в зависимости от погоды (рис. 24.8). Таким образом, птицы, по всей вероятности, извлекают гораздо больше пользы от того, что сначала поют, а потом начинают кормиться, чем если бы они поступали наоборот.
Чтобы судить о том, является ли одна последовательность действий лучше, чем другая, нам нужно иметь какую-то общую «валюту», с помощью которой можно было бы сравнить эти последовательности. Нам также необходимо иметь определенный критерий, с помощью которого можно было бы измерять относительную ценность различных видов поведения. Каждый вид активности характеризуется затратами и выигрышами, связанными с ее реализацией. В этой главе мы рассмотрим вопрос о том, как можно точно оценить эти затраты и выигрыши, однако пока мы ограничиваемся общими рассуждениями. В конечном счете, какая разница, приводит ли данная конкретная активность к увеличению или снижению общей приспособленности животного? Большая синица, которая поет на рассвете, повышает свою приспособленность тем, что не допускает на участок кон-
|
| Рис. 24.8. Зависимость освещенности мест, где кормится большая синица, от времени суток (в солнечную и пасмурную погоду). Р и S обозначают соответственно уровни освещенности, при которых рентабельность пищевого поведения и эффективность поиска пищи достигают (по данным лабораторных измерений) 95% своей максимальной величины. (По Kacelnik, 1979.) |
курентов, но в то же время ее приспособленность уменьшается за счет того, что она откладывает на потом добывание пищи, особенно если она потеряла в весе за ночь. Такое повышение приспособленности удобно называть выигрышем (прибылью), а понижение - затратами. Так, например, при добывании пищи животное выигрывает, приобретая энергию, но может потерпеть убыток (понести затраты), пренебрегая защитой своего участка или становясь более уязвимым для хищника. Одно из преимуществ оценки поведения животного с точки зрения затрат и выигрыша состоит в том, что оно позволяет нам сравнивать относительные ценности совершенно разных активностей. Различные виды пищедобывательного поведения можно сравнить с точки зрения приобретенных неэнергетических компонентов пищи и энергии, а различные типы защиты территории можно сравнивать по их эффективности в противостоянии пришельцам. Однако при сравнении пище-добывания и поведения защиты мы должны определять, намного ли каждая из этих активностей увеличивает или уменьшает общую приспособленность животного. Приспособленность - это единственная общая «валюта», с помощью которой мы можем проводить сравнения различных видов поведения. В конечном счете именно с помощью этой валюты природа производит такое сравнение, поскольку приспособленность определяется через действие естественного отбора.
Представляется важным не только иметь возможность определить, к каким последствиям могут приводить различные виды активности в целом, но еще и оценить их с точки зрения некоторого критерия. Так, например, вполне возможно, что при рассмотрении кандидатур на должность преподавателя университета мы захотим сравнить кандидатов по обычным показателям, таким, как педагогические способности и его потенциальные возможности как исследователя. Кроме того, нам нужен еще и критерий, с помощью которого мы могли бы решить, кому из кандидатов отдать предпочтение: хорошему преподавателю, но плохому исследователю или слабому преподавателю, но хорошему исследователю (McFarland, 1976; 1977). Подобным же образом при оценке поведения животного нам необходим критерий оптимальности, который позволял бы судить о том, будет ли при определенных условиях один вид активности лучше, чем другой. Хотелось бы надеяться, что в конечном счете мы сможем определить критерий оптимальности с точки зрения общей приспособленности животного. Однако для того чтобы сделать это грамотно, потребовались бы обширные знания о состоянии животного и об условиях, в которых оно находится. В следующем разделе мы рассмотрим возможные подходы к получению таких знаний.
Функции затрат
Мы уже видели, что животным часто приходится выбирать между альтернативными последовательностями действий, характеризующихся определенным балансом затрат и выгод. Наилучшим вариантом поведения животного будет тот, который обеспечивает наименьшие затраты или наибольшие выгоды. Подходя с эволюционных позиций, мы можем ожидать, что хорошо организованные животные должны вести себя так, чтобы максимизировать свою общую приспособленность. Таким образом, концепция затрат и выгод оказывается связанной с концепцией приспособленности.
Животные располагают большим разнообразием способов осуществления затрат и получения выгод, которые соответствуют давлениям естественного отбора. Естественный отбор действует на животное в целом; однако анализируя затраты и выигрыши от той или иной формы поведения, мы должны разделять затраты (уменьшение приспособленности) и выигрыши (увеличение приспособленности), которые могут быть приписаны каждому виду внутреннего состояния и поведения животного. Совокупность характеристик животного, представленная в таком виде, называется функцией затрат. Ее можно определить как характеристику мгновенного уровня риска, которому подвергается животное (и репродуктивных преимуществ, которые оно имеет), когда оно находится в определенном внутреннем состоянии и осуществляет определенную деятельность в определенных условиях среды (McFarland, 1977).
Предположим, например, что мы изучаем поведение серебристой чайки (Larus argentatus), насиживающей яйца в гнезде. Обычно оба родителя насиживают яйца по очереди, и если только гнездо останется без внимания, его могут быстро разграбить соседи. Сидящая птица не оставляет гнезда до тех пор, пока ее не сменит супруг или не спугнет какой-либо хищник, например лисица или человек. Обычно птица покидает гнездо для добывания пищи и возвращается к нему через несколько часов. Однако иногда ее возвращение может задержаться из-за какой-либо неудачи, несчастного случая, а порой и из-за того, что ее может поймать экспериментатор. Что в таком случае должна делать сидящая на гнезде птица? С одной стороны, ее супруг может вернуться в любое время, но с другой - по мере течения времени у нее усиливается чувство голода. В конце концов эта птица будет вынуждена покинуть гнездо, чтобы добыть пищу. Поскольку серебристые чайки способны размножаться круглый год, с генетической точки зрения представляется невыгодным подвергать опасности жизнь особи из-за единичной кладки яиц. Вопрос в том, в какой именно момент птица должна решиться покинуть гнездо?
Если «наседка» улетает с гнезда слишком рано, то она подвергает опасности свою кладку без особой на то необходимости, потому что ее супруг, быть может, просто ненадолго задерживается. Если же птица покинет гнездо слишком поздно, то она необоснованно подвергает опасности свою жизнь, поскольку она может не найти пищу так быстро, как это бывает обычно. Для того чтобы точно определить, когда же птица должна улететь с кладки, необходимо количественно оценить степени риска в каждой из этих альтернативных ситуаций. Такую оценку эффективно осуществляет природа, создавая животное, которое будет принимать самое подходящее в этой ситуации решение.
Чтобы изучить эту проблему, необходимо в полевых условиях измерить степень риска, которому подвергаются животные, занятые различными видами активности. Так, например, в случае насиживающих яйца серебристых чаек нам необходимо знать степень риска, которому подвергнутся яйца, если останутся в гнезде без присмотра, и степень риска, которому подвергнется птица, если будет продолжать сидеть на гнезде без пищи. Были предприняты попытки в самых различных экспериментах оценить затраты и выгоды, присущие поведению насиживания. Так, например, Дрент (Drent, 1970) изучал поведение насиживания яиц у серебристых чаек на о. Схирмонниког (Западно-Фризские о-ва). Он обнаружил, что
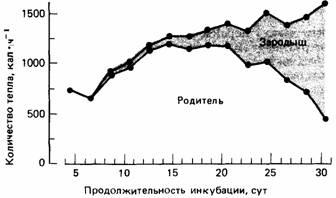
Рис. 24.9.Энергетическая цена инкубации яиц у серебристых чаек, вычисленная для всей кладки. Тепло, которое производит сам зародыш, изображено в виде затушеванной области. Таким образом, верхняя кривая отражает общее количество необходимой энергии, а нижняя - энергозатраты родителей. (По Drent, 1970.)
20% яиц уничтожается хищниками, а 12% погибает от того, что не соблюдаются физические условия, необходимые для их инкубации. Если человек не тревожит птиц, они проводят на яйцах около 98% времени. Благодаря этому яйца защищены от переменчивых воздействий внешней среды, а птица имеет возможность регулировать температуру яиц. Главная проблема, с которой сталкивается птица, насиживающая яйца, - это регулирование температуры собственного тела. Холодной ночью ей приходится продуцировать тепла столько, чтобы его хватило для согревания себя и яиц, тогда как в полуденную жару ей приходится дышать очень часто, чтобы несколько охладить себя. Дрент присоединял датчики к гнезду и яйцам и с помощью следящей (monitoring) аппаратуры смог детально исследовать роль факторов, влияющих на развитие зародышей чаек. В самом начале инкубации температура яиц полностью зависит от родителей, но по мере развития зародыш начинает продуцировать метаболическое тепло (рис. 24.9). Однако зародыши не способны к терморегуляции, и поэтому, если оставить яйца открытыми, они будут постоянно охлаждаться. У зародыша нет компенсаторной теплопродукции, а вот только что вылупившийся птенец реагирует на снижение температуры воздуха повышением теплопродукции. Таким образом, птенец в какой-то степени обладает температурным гомеостазом.
В отсутствие родителей зародыш рискует погибнуть либо от перегревания, либо от переохлаждения в зависимости от того, какая стоит погода. Однако более важным фактором является общее количество тепла, которое получает зародыш к моменту его вылупления. Скорость развития зародыша зависит от интенсивности его метаболизма, на которую в свою очередь влияет температура внутри яйца. Если эта температура низкая, то зародыш развивается медленно. Во время инкубации яйцо постоянно теряет в массе за счет испарения воды (рис. 24.10). Такая потеря
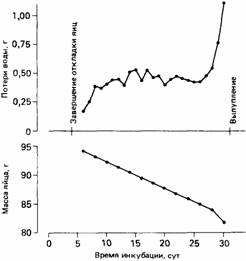
Рис. 24.10.Потери воды (верхний график) яйцами серебристой чайки, измеренные по изменениям их массы (нижний график). По мере потери воды воздушное пространство внутри яйца увеличивается, однако слишком большая потеря воды приводит к обезвоживанию зародыша. (По Drent, 1970.)
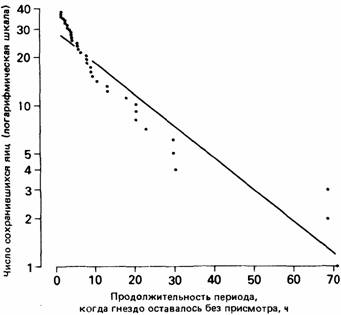
| Рис. 24.11. Сохранность яиц серебристой чайки, оставшихся на виду у хищников в отсутствие владельпев гнезда. (Данные из работы Sibly, McCleery, 1985.) |
воды приводит к тому, что в одном конце яйца образуется воздушное пространство, необходимое для дыхания зародыша в период, непосредственно предшествующий вылуплению. Зародыш не должен развиваться слишком быстро, иначе это воздушное пространство не сформируется как следует к тому моменту, когда птенец будет готов к вылуплению. Зародыш не должен развиваться и слишком медленно, иначе яйцо потеряет так много воды, что наступит дегидратация зародыша. Вылупление должно произойти в тот момент, когда яйцо потеряет строго определенное количество воды, а зародыш к этому времени будет достаточно развит, чтобы покинуть яйцо. Не имеет столь большого значения кратковременное охлаждение яиц, важно, чтобы за это время их температура не упала слишком низко. В теплый день яйца подстерегает другая опасность: они могут перегреться, если останутся долго открытыми, но обычно яйца, оставленные открытыми на час или два, с физиологической точки зрения практически не страдают. Таким образом, серебристые чайки во время насиживания яиц вполне могут пережить случайные тревоги, которые вынуждают их покидать гнезда. Однако если чайки будут потревожены не один раз, то средняя скорость притока тепла к яйцам может резко упасть ниже того уровня, который требуется для успешного вылупления.
У зародышей серебристой чайки в два раза больше шансов погибнуть от хищников, чем от нарушения их развития. Сидящим на кладке птицам приходится покидать свои гнезда при появлении некоторых хищников, а также человека. В отсутствие родителей яйца могут стать добычей соседних чаек (чаек-каннибалов), крыс, ежей, ворон или других хищников. Уровень риска, которому подвергаются яйца, может быть рассчитан на основе полевых наблюдений за судьбой оставшихся без присмотра яиц или в специальных экспериментах, когда родителей удаляют из гнезда (рис. 24.11).
Чайка, сидящая в гнезде и ожидающая возвращения своего партнера, не может покинуть гнездо, не подвергая опасности яйца. Как мы уже видели, риск, которому подвергаются яйца, можно количественно оценить как часть общей функции затрат. С одной стороны, имеются затраты как результат прекращения насиживания, поскольку возникает риск, что из яиц не вылупятся птенцы, потому что их съедят хищники или они погибнут от непогоды. С другой стороны, имеются затраты, связанные с насиживанием, которые склады-

Рис. 24.12. Эти весы (слева) для взвешивания гнезда можно расположить в ямке, выкопанной под гнездом серебристой чайки. Затем гнездо укладывают на платформу весов (справа). Поведение птиц на гнезде при этом не изменяется, а изменения массы гнезда можно регистрировать с помощью соответствующего электронного устройства. (Фотография Robin McCleery.)
ваются из риска быть пойманным хищником и из риска погибнуть от истощения.
Указанные аспекты функции затрат также могут быть исследованы экспериментально. Например, массу тела насиживающей птицы можно измерить с помощью специальных весов, помещенных в яму, выкопанную в земле под гнездом (рис. 24.12). Количество жира у птицы можно вычислить исходя из соотношения ее массы и размеров костей скелета, которые измеряются у пойманной птицы перед началом эксперимента в процессе ее мечения. Количество пищи, которое серебристая чайка получила за время кормежки, можно оценить по изменению ее массы, измеренной на тех же весах, помещенных под гнездом, перед и после ее путешествия за пищей. Качество пищи можно оценить путем анализа фекалий, собранных вокруг гнезда, и по результатам наблюдений за птицей во время добывания ею пищи. Серебристые чайки могут летать за пищей на расстояния в несколько миль. Поэтому при проведении наблюдений за их пищедобывательной активностью некоторых птиц снабжали радиопередатчиками (рис. 24.13) и прослеживали их маршрут с помощью радиоприемника с направленной антенной (рис. 24.14). Таким образом, используя различную аппаратуру, иногда удается получить достаточно точные оценки затрат и выигрышей животных, ведущих обычный образ жизни в естественных условиях обитания (Sibly, McCleery, 1985).
Вообще говоря, функция затрат состоит из двух главных частей. Первую часть составляют затраты, связанные с активностью, осуществляемой в данное время, а вторую - затраты, связанные с тем, что другие активности в это время не осуществляются. Например, чайка, сидящая на яйцах, несет затраты, связанные с их насиживанием. В них входят физиологические затраты, связанные с поддержанием определенной температуры яиц, и поведенческие затраты, связанные с риском быть пойманным хищником во время насиживания. Кроме того, некоторые затраты обусловлены тем, что насиживание несовместимо с другими видами поведения, обычно связанными с иными мотивациями животного. Так, например, во время насиживания у птиц неизбежно усиливается чувство голода. А голод влечет за собой риск гибели от истощения, поскольку в тот момент, когда животное решит приступить к добыванию пищи, оно может не найти ее. Как мы увидим далее, такой анализ поведения обязательно требует знания особенностей экологии данного животного. Стараясь понять, каким образом животные учитывают затраты и выигрыши, обусловленные особенностями среды их обитания, иногда бывает полезно провести аналогию между экономическими расчетами человека и подобным поведением у животных.

Рис. 24.13.Серебристая чайка с прикрепленным к ней радиопередатчиком. (Фотография Robin
McCleery.)
Дата добавления: 2016-06-18; просмотров: 3454;
Поиск по сайту
Узнать еще
- I.1.9 ЯВЛЕНИЯ ПЕРЕНОСА В ГАЗАХ
- I.3.9 ТЕПЛОВЫЕ ЯВЛЕНИЯ В СРЕДАХ ПРИ МЕХАНИЧЕСКОМ ВОЗДЕЙСТВИИ
- II. Выявление причин появления события.
- А. Причины появления и способа уменьшения нелинейных искажений
- Адаптация растений и животных к световому режиму.
- Адгезионные явления
- Анализ внешних факторов с целью выявления причин кризиса.
- Аналогия явления переноса субстанций. Законы молекулярного переноса

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
