ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ ПО НЕРВНЫМ ВОЛОКНАМ
Как известно, потенциалы могут быть локальными, способными распространяться с декрементом (затуханием) всего на 1-2 мм. и импульсными (ПД), которые распространяются без декремента по всей длине нервного или мышечного волокна - до нескольких десятков сантиметров: например, от мотонейронов спинного мозга до мышечных волокон конечностей с учетом и длины самих конечностей. Локальные потенциалы могут возникать в любой клетке, способной генерировать ПД при действии подпорогового раздражения, а также в структурах нервной ткани, не имеющих быстрых потенциалзависимых каналов. Таковыми являются постсинаптические мембраны, мембраны сенсорных рецепторов, часть мембраны тела нейрона и дендрита, клетки нейроглии.
А. Механизм проведения локального потенциала.Локальные потенциалы (препотенциал, рецепторный потенциал, возбуждающий постсинаптическин потенциал) изменяют мембранный ПП, как правило, в сторону деполяризации в результате входа в клетку ионов N3*. В результате этого между участком волокна, в котором возник локальный потенциал, и соседним участком мембраны формируется разность потенциалов, вызывающая передвижение ионов согласно электрическому градиенту. В частности, ионы Nа+ на наружной поверхности мембраны начинают перемещаться в сторону того участка, в котором первично возник локальный потенциал, при этом положительный потенциал наружной поверхности мембраны соседнего участка уменьшается. Аналогичные изменения наблюдаются на внутренней поверхности мембраны, где вошедшие в клетку ионы Nа+ движутся в обратном направлении, что также ведет к уменьшению отрицательного потенциала соседнего участка. В итоге поляризации соседнего участка мембрана уменьшится.Возникшая частичная деполяризация соседнего участка клеточной мембраны вызывает при достижении 50% Екр открытие рядом расположенных потенциалчувствительных ионных каналов клеточной мембраны (см. раздел 2.6.4), что обеспечивает ионный ток в этом месте и, как следствие, частичную деполяризацию мембраны - локальный потенциал. Последний также действует на соседний участок мембраны и т.д., при этом главное направление движения ионов (Nа+ - внутрь клетки, ионов К* - из клетки), т. е. перпендикулярно клеточной мембране. Если деполяризация соседнего участка мембраны также не достигает критического уровня, то она не обеспечивает возникновения ПД. При этом быстро развивается инактивация Na-каналов, поэтому деполяризация быстро сменяется рсполяризацией вследствие выхода ионов К+ из клетки по неуправляемым каналам (каналы утечки ионов), в результате чего локальный потенциал затухает.
Подобный механизм распространения локального потенциала наблюдается только в тех мембранах, которые содержат потенциал-чувствительные ионные каналы.
Если возникшая деполяризация мембраны не сопровождается изменением проницаемости потенциалзависимых натриевых, кальциевых и калиевых каналов, такую деполяризацию называют электротонической(физической).
Электротоническое распространение возбуждения характерно для фрагментов мембран возбудимых клеток, где нет потенциалзависимых ионных каналов, и ионы движутся только вдоль мембраны волокна. Такими участками являются, например, мембраны дендритов большинства нервных клеток, межпере-хватные промежутки в миелиновых нервных волокнах. Если распространяющееся локальное возбуждение достигает участков мембраны, способных генерировать ПД (перехваты Ранвье, ак-сонный холмик нейрона), и их амплитуда выходит на критический уровень деполяризации, формируется ПД, распространяющийся по всей длине волокна.
Эффективность электротонического распространения биопотенциалов зависит от физических свойств нервного волокна: сопротивления, емкости мембраны, сопротивления цитоплазмы.Электротоническое проведение в нервном волокне улучшается при увеличении его диаметра, что связано с уменьшением сопротивления цитоплазмы, а также при миелинизации волокна, увеличивающей сопротивление мембраны. Проводимость нервного волокна для локального потенциала характеризует постоянная длины мембраны Lm - расстояние, на которое может электротонически распространиться биопотенциал, пока его амплитуда не уменьшится до 37% от исходной величины. Постоянная длины мембраны тонких безмиелиновых волокон не превышает 1 мм, толстых миелиновых волокон достигает 5 мм.
Передача информации на большие расстояния в пределах нервной системы осуществляется с помощью нервных импульсов (ПД) по аксонам нейронов.
Б. Механизм проведения потенциала действия. Обязательным условием проведения нервного импульса является наличие на всем протяжении или в ограниченных, но повторяющихся участках волокна быстрых потенциалзависимых (потенциалчув-ствительных) каналов, ответственных за формирование ПД. В распространении ПД можно выделить два этапа: этап электротонического проведения, обусловленного физическими свойствами нервного волокна, и этап генерации ПД в новом участке на пути его движения. В зависимости от расположения и концентрации ионных каналов в мембране волокна возможно два типа проведения ПД: непрерывный и сальтаторный (скачкообразный).
1. Непрерывное проведение нервного импульса (ПД) осуществляется в безмиелиновых волокнах типа С, что объясняется равномерным распределением потенциалчувствительных ионных каналов, участвующих в генерации ПД. Возникший ПД обеспечивает открытие потенциалзависимых Ка-каналов на соседнем участке мембраны нервного волокна и движение ионов Na+ внутрь волокна и вдоль волокна, т.е. электротонически, что обеспечивает развитие критического уровня деполяризации на соседнем участке нервного волокна и возникновение нового ПД. Постоянная длины мембраны (Lm) безмиелиновых волокон составляет примерно 0,1-1,0 мм, т.е. эта величина во много раз больше расстояния между отдельными каналами, что обеспечивает высокую надежность проведения ПД, способного деполяризовать мембрану до критического уровня и обеспечить генерацию нового ПД (рис. 4.1). Следует также отметить, что чисто электротонический этап распространения ПД (вдоль мембраны) в безмиелиновом волокне предельно мал и наблюдается только до достижения деполяризации мембраны 50% ЕКр. Далее включается перпендикулярное перемещение ионов за счет активации ионных каналов, при этом в каждом участке мембраны ПД возникает заново, поэтому он проводится без снижения амплитуды - без декремента.
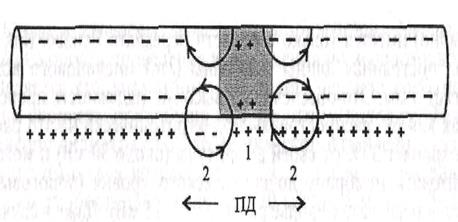
| Рис. 4.1. Непрерывное распространение ПД в нервном волокне. 1 - возникновение ПД в нервном волокне и распространение ПД в обе стороны от места возникновения. В области деполяризации мембраны (I) преобладает входящий в клетку натриевый ток; 2 - соседняя область, в которой локальный ток от области ПД вызывает деполяризацию до критического уровня | |
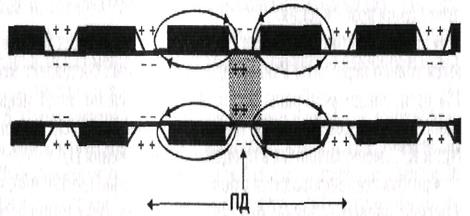
| Рис. 4.2. Сальтаторное распространение ПД в миелиновых нервных волокнах. Возникновение ПД в перехвате Ранвье среднего участка волокна и распространение ПД в обе стороны от места возникновения. Стрелками показаны токи на электротоническом этапе распространения ПД. В области перехватов Na+ движется в клетку, К+ - из клетки | |
Непрерывное распространение нервного импульса идет через генерацию новых ПД по эстафете, когда каждый возник ший импульс является раздражителем для соседнего участка нервного волокна и обеспечивает возникновение нового ПД.
2. Сальтаторное проведение нервного импульса (ПД) осуществляется в миелиновых волокнах, так как у них потенциалчувстви-тельные ионные каналы локализованы только в участках мембраны перехватов Ранвье, где их плотность достигает 12 000 на 1 мкм2. В области миелиновых муфт (межузловые сегменты), обладающих высокими изолирующими свойствами, потенциалчувствительных каналов почти нет, вследствие чего мембрана осевого цилиндра там практически невозбудима. Поэтому ПД, возникший в одном перехвате Ранвье, электротонически (продольно, без участия ионных каналов) распространяется до соседнего перехвата и деполяризует мембрану до критического уровня, что ведет к возникновению нового ПД, т.е. возбуждение проводится скачкообразно (рис. 4.2). Движение ионов Nа+ внутрь миелинового волокна (перпендикулярно) имеется только в области перехвата Ранвье. Как отмечалось, постоянная длины мембраны (Ат) миелинового волокна достигает 5 мм. Это обеспечивает высокую надежность проведения ПД, так как он, распространяясь электротонически на это расстояние, сохраняет 37% от своей амплитуды (около 30 мВ) и может деполяризовать мембрану до критического уровня (пороговый потенциал в перехватах Ранвье равен около 15 мБ). Даже в случае повреждения ближайших перехватов Ранвье на пути следования ПД он может электротонически возбудить 2-4-й и даже 5-й перехваты. Сальтаторное проведение нервных импульсов является эволюционно более поздним механизмом, возникшим впервые у позвоночных в связи с миелинизацией нервных волокон. Оно имеет два важных преимущества по сравнению с непрерывным механизмом проведения возбуждения:
• более экономично в энергетическом плане, так как возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% от площади мембраны волокна, следовательно, надо меньше энергии для восстановления трансмембранных градиентов ионов Na+и К+, уменьшающихся в процессе возникновения ПД;
• возбуждение проводится с большей скоростью (до 120 м/с), чем в безмиелиновых волокнах (0,5-2,0 м/с), так как электротоническое распространение ПД в области миелиновых муфт происходит значительно быстрее, чем в результате непрерывной генерации ПД в безмиелиновых волокнах. В связи с этим миелиновые волокна в нервной системе сформировались там, где необходима наиболее быстрая регуляция функций. В миелиновых волокнах ПД как бы перескакивает от одного перехвата Ранвье к другому.
В. Закономерности проведения возбуждении по нервным волокнам.
1. Двустороннее проведение возбуждения доказывается регистрацией нервных импульсов с двух сторон от места нанесения раздражения на нерв, способного вызвать возбуждение (рис. 4.1). В условиях целого организма двустороннее проведение наблюдается в аксонном холмике нейрона. Возникающий в этом месте ПД переходит не только на аксон, но и на тело нейрона.
2. Изолированное проведение возбуждения в отдельных нервных волокнах. Обычно оно не передается с одного нервного волокна на другое. Это обусловлено тем, что петли тока в межклеточной жидкости ствола, имеющей низкое сопротивление, почти не проникают в невозбужденные волокна нерва вследствие высокого сопротивления их оболочек. Изолированное проведение импульсов по нервным волокнам обеспечивает высокую точность регуляторной деятельности ЦНС на другие нервные клетки и клетки-эффекторы рабочего органа.
3. Большая скорость проведения возбуждения, достигающая 120м/с. Скорость проведения возбуждения по нервному волокну определяется его диаметром: чем толще нервное волокно, тем больше скорость проведения ПД по этому волокну. Так, при увеличении диаметра от 12 до 22 мкм скорость распространения нервного импульса возрастает от 70 до 120 м/с, при диаметре 8-12 мкм скорость составляет 40-70 м/с, 4-8 мкм - 15-40 м/с, 1 -4 мкм — 5-15 м/с.
4. Неутомляемость нервного волокна. Н.Е.Введенский (1883) обнаружил, что нерв сохраняет способность к проведению возбуждения в течение 6-8 ч непрерывного раздражения. Это обусловлено тем, что при проведении ПД по нервным волокнам используется 1/1 000 000 часть запасов трансмембранных ионных градиентов и, следовательно, нужны небольшие количества АТФ для восстановления ионных градиентов. Расход энергии в нерве примерно в 16 раз меньше, чем на соответствующую единицу массы в целом организме в условиях покоя.
5. Возможность функционального блока проведения возбуждения при морфологической целостности нервных волокон. Н. Е. Введенский (1901) показал, что действие различных альтерирующих агентов на нерв вызывает состояние парабиоза, в результате чего нервный импульс не проходит через этот участок нерва. Причиной блока являются специфические расстройства работы ионных каналов мембраны под влиянием альтерирующих агентов. Для создания блока протяженность парабиотического участка должна превысить постоянную длины мембраны, иначе ПД может распространиться через этот участок электротонически. Нарушение физиологической непрерывности нервных волокон можно наблюдать при действии на нерв анестетиков, различных ядов, растворов солей, новокаина, при гипоксии, охлаждении. Временная блокада проведения возбуждения широко применяется в клинической практике.
Дата добавления: 2021-12-14; просмотров: 534;
Поиск по сайту
Узнать еще
- Автоматическими регуляторами возбуждения (АРВ)
- Агротехнические требования на проведение механизированных работ
- Анализ исходных данных. Проведение оценки значимости экологической ситуации
- Анкетирование и проведение интервью
- Аритмии вследствие нарушения возбудимости и проведения возбуждения в сердечной мышце.
- Атрибуции при переносе возбуждения
- Борьба с АСПО предусматривает проведение работ по двум направлениям: по предупреждению образования отложений и удалению уже образовавшихся.
- В качестве примера рассмотрим проведение занятия по ознакомлению с деньгами.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
