Использование энергии света
Способность использовать энергию света обуславливается наличием у большинства фототрофных микроорганизмов специфических пигментов - бактериохлорофиллов и каротиноидов.
Световая энергии улавливается системой поглощающих свет пигментов и передается в реакционный центр, который содержит молекулы хлорофилла. Когда свет падает на молекулу хлорофилла, она переходит в возбужденное состояние вследствие перехода одного из электронов на более высокий энергетический уровень. Переходя по цепи переноса, электрон отдает свою энергию системе АДФ-АТФ, в результате чего энергия света трансформируется в энергию макроэргической связи молекулы АТФ, т.е. происходит фотосинтетическое фосфорилирование, которое бывает циклическое и нециклическое.
В случае циклического процесса электрон возвращается к исходному донору, отдавая полученную им энергию в процессе перехода через ряд переносчиков по термодинамическому закону, одним из таких переносчиков является цитохром с, обеспечивающий фосфорилирование АДФ. При нециклическом фосфорилировании возбужденный электрон передается на НАД+ с образованием НАДН2.
У фотосинтетических бактерий донорами водорода для реакций синтезе могут быть как неорганические, так и органические вещества. Фотолитоавтотрофы используют в качестве донора водорода Н2S:
СО2 + Н2S → (СН2О) + Н2О + 2 S (3.4)
Такой вид фотосинтеза называется фоторедукцией. Основное отличие бактериальной фоторедукции от фотосинтеза зеленых растений и водорослей следующие: бактериальный фотосинтез протекает в анаэробных условиях и не сопровождается выделением кислорода; донором водорода для восстановления СО2 в большинстве случаев является сероводород, а не вода.
Микроскопические водоросли и цианобактерии (как и высшие растения) фотосинтез осуществляют с выделением О2, донором электронов (водорода) служит вода:
СО2 + Н2О → (СН2О) + О2 (3.5)
Брожение
Брожение – это способ получения энергии без участия атмосферного кислорода в результате окислительно-восстановительных реакций, в которых органические вещества функционируют как доноры и как акцепторы водорода.
Сбраживаться могут не все вещества, а только такие, которые имеют неполностью окисленные (или восстановленные) атомы углерода и поэтому способны подвергаться сопряженному процессу окисления–восстановления, приводящему к выделению энергии. В процессах брожения расщепляются вещества различной степени сложности. Наиболее доступным органическим субстратом для процессов брожения являются гексозы, в частности глюкоза.
Брожения более сложных субстратов осуществляется после предварительного расщепления их до глюкозы или продуктов ее превращения. Некоторые микроорганизмы способны извлекать энергию при сбраживании пентоз, жирных кислот, аминокислот. У микроорганизмов есть несколько серий реакций, ведущих к выработке энергии путем расщепления гексоз в условиях анаэробиоза.
Гексозодифосфатный путь. Это наиболее распространенный путь. Назван он так по основному промежуточному продукту этого пути – дважды фосфорилированному сахару - дифосфофруктозе (фруктозо-1,6-фосфат). Этот путь носит название гликолиза (греч. glikos– сладкий, lisis – расщепление). По имени ученых, расшифровавших основные реакции этого пути, его называют также путь Эмбдена-Мейергофа-Парнаса (ЭМП-путь). В процессе гликолиза синтез АТФ происходит на уровне превращения 1,3-дифосфоглицериновой кислоты в 3-фосфо-глицериновую кислоту и фосфоенолпировиноградной кислоты в пировиноградную кислоту (пируват). В результате расщепления глюкозы в процессе гликолиза расходуется две, а синтезируется четыре молекулы АТФ. Таким образом, общий выход составляет двемолекулы АТФ и две молекулы НАДН2 (рисунок 3.2).
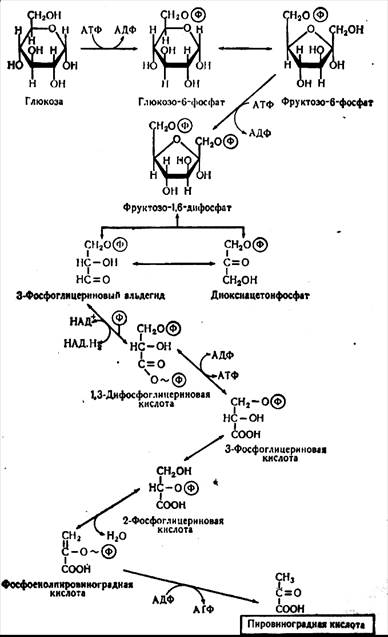
Рисунок 3.2 – Схема гликолитического пути расщепления глюкозы
Гексозомонофосфатный путь (ГМФ-путь). Система реакций этого пути позволяет использовать в качестве энергетического материала, не только гексозы, но и пентозы. Поэтому путь называется также пентозофосфатный окислительный путь (ПФ-путь). Этот путь необходим также для синтеза рибоз, необходимых для нуклеиновых кислот и других соединений. Эта система реакций носит также название путь Варбурга-Диккенса-Хорекера (рисунок 3.3).
Первая часть реакций этого пути представляет окислительный процесс, начинающийся также с фосфорилирования глюкозы, которая затем подвергается дважды дегидрированию и один раз декарбоксилированию. В результате образуется центральный метаболит этого пути – рибулозо-6-фосфат. Дальнейшая серия реакций представляет собой взаимопревращения углеродных соединений. Особенностью распада углеводов по ГМФ-пути является образование НАДФН2, а не НАДН2, как при гликолизе.
У анаэробных микроорганизмов этот путь обычно функционирует параллельно с гликолизом, так как ГМФ-путь сам по себе в анаэробных условиях не приводит к синтезу АТФ. Только у аэробных микроорганизмов этот путь может быть источником энергии после того, как оторванный с помощью НАДФ+, водород поступает в дыхательную цепь.
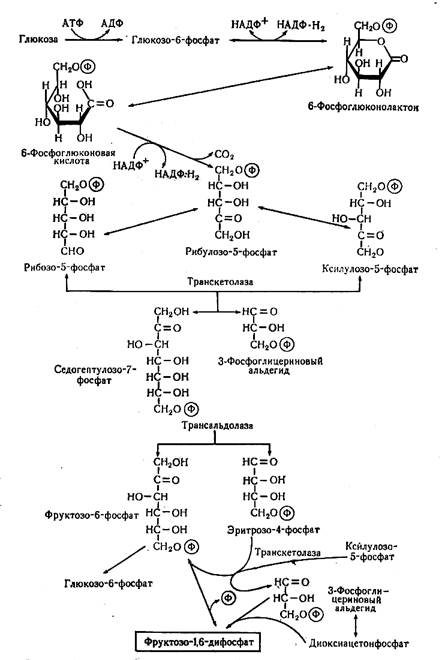
Рисунок 3.3 – Пентозофосфатный окислительный путь
Кетодезоксифосфоглюконатный путь (КДФГ-путь). Этот путь обнаруживается только у микроорганизмов и представляет собой модификацию гликолиза и ГМФ-пути. По имени ученых, открывших путь, он называется путь Энтнера-Дудорова (рисунок 3.4). Этот путь используется микроорганизмами для получения ПВК более коротким путем. Если при гликолизе для получения ПВК надо пройти девять этапов, то при КДФГ-пути достаточно четырех этапов. У некоторых микроорганизмов этот путь может быть единственным для получения энергии при усвоении сахаров в анаэробных условиях. КДФГ-путь также начинается с фосфорилирования глюкозы и образования АДФ из АТФ. Затем глюкозо-6-фосфат превращается в 6-фосфо-глюконовую кислоту, от которой отщепляется вода и образуется 2-кето-3-дезокси-6-фосфо-глюконовая кислота. Это соединение расщепляется специфической альдолазой на ПВК и 3-фос-фоглицериновый альдегид, окисляющийся в ПВК. При расщеплении глюкозы КДФГ-путем образуется по одной молекуле АТФ, НАДН2 иНАДФН2.
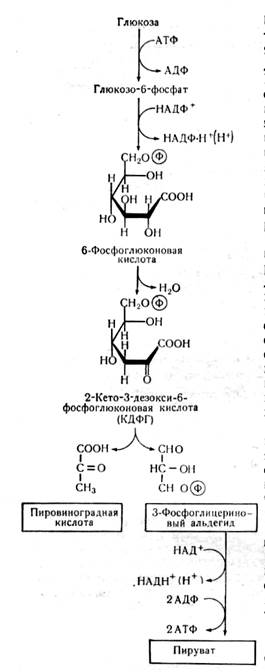
Рисунок 3.4 – Кетодезоксифосфоглюконатный путь
Для анаэробных микроорганизмов представленные процессы являются основной возможностью получения энергии при усвоении сахаров. В зависимости от наличия специфических для каждого вида микроорганизмов ферментов в анаэробных условиях происходит перенос водорода от НАДН2 на ПВК или субстраты, образуемые из ПВК. В результате этого получаются различные восстановленные соединения, называемые продуктами брожения, а соответственно им называются процессы брожения. Различают несколько типов брожения – молочнокислое, маслянокислое, спиртовое и др., которые вызываются соответствующими микроорганизмами.
Молочнокислое брожение. Это анаэробное превращение углеводов с образованием молочной кислоты. Существуют два вида молочнокислого брожения - гомоферментативное и гетероферментативное. Вызываются эти виды брожения соответственно гомоферментативными и гетероферментативными молочнокислыми бактериями (МКБ).
Гомоферментативное молочнокислое брожение. Сначала расщепление глюкозы идет по ЭМП пути (гликолиз). Гомоферментативные МКБ синтезируют фермент лактатдегидрогеназу (ЛДГ), который использует ПВК в качестве акцептора водорода и восстанавливают ее до молочной кислоты:
ЭМП-путь ЛДГ
С6Н12О6 → 2 СН3СОСООН + 2 НАДН2 → 2 СН3СНОНСООН + НАД+ (3.6)
Гетероферментативное молочнокислое брожение. Гетероферментативные МКБ не имеют альдолазы и триозофосфатизомеразы (основные ферменты, необходимые для расщепления углеводов по ЭМП-пути), поэтому эти микроорганизмы метаболизируют глюкозу по ГМФ-пути. При гетероферментативном молочнокислом брожении одновременно с образованием молочной кислоты происходит накопление этилового спирта, СО2, уксусной кислоты и других продуктов. Образование конечных продуктов зависит от вида микроорганизма, условий культивирования (рН, температура, степень аэрации) и других факторов. Например:
2 С6Н12О6→СН3СНОНСООН+СООНСН2СН2СООН+СН3СООН+СН3СН2ОН+Н2 (3.7)
Молочнокислые бактерии – это бактерии, способные образовывать в качестве главного продукта брожения молочную кислоту. В группу МКБ объединены различные по систематическому положению микроорганизмы: молочнокислые кокки(представители родов Lactococcus, Pediococcus, Aerococcus, Leuconostoc), молочнокислые палочки (род Lactobacillus). МКБ широко распространены в природе. Они встречаются на различных растениях, в почве (в верхних слоях), в ризосфере растений, в желудочном тракте теплокровных животных и человека, в силосе, муке, овощах, сыром молоке, молочных продуктах, различных квашеных продуктах. На основе живых культур лактобактерий изготовлены многочисленные пробиотические препараты, большинство из них предназначено для коррекции микрофлоры организма человека.
Спиртовое брожение. Спиртовое брожение вызывают различные дрожжи и некоторые бактерии. Для большинства микроорганизмов спирт является побочным продуктом и только для дрожжей рода Saccharomyces – это главный конечный продукт брожения.
Перед началом спиртового брожения олигосахара вначале гидролизуются соответствующими ферментами дрожжей до гексоз. Затем гликолитическим путем осуществляется расщепление гексоз и образование ПВК. Под действием пируватдекарбоксилазы (ПДК) микроорганизмов от ПВК отщепляется СО2 и образуется ацетальдегид:
ЭМП-путь ПДК
С6Н12О6 → 2 СН3СОСООН → 2 СН3СНО + 2 СО2 (3.8)
Ацетальдегид служит конечным акцептором водорода. Он восстанавливается в этанол при участии фермента алкогольдегидрогеназы (АДГ) Брожение предполагает строгое равновесие процессов окисления и восстановления. Поэтому НАД, восстановленный на одном из этапов брожения, должен окисляться на другом этапе. Окисление НАДН2 происходит одновременно с восстановлением ацетальдегида в этанол:
АДГ
2 СН3СНО + 2 НАДН2 → 2 СН3СН2ОН + 2НАД+ (3.9)
Таким образом, при спиртовом брожении основными продуктами превращения сахаров являются этанол и СО2:
С6Н12О6 → 2 СН3СН2ОН + 2 СО2 (3.10)
Спиртовое брожение протекает наиболее интенсивно в кислой среде (рН 4,0…4,5), при температуре 30оС и концентрации сахара 10…15%. Повышенная концентрация сахара приводит к замедлению, а затем прекращению процесса.
Ход брожения может заметно меняться в зависимости от конкретных условий. Если в культуру бродящих дрожжей добавить бисульфит натрия, то он связывает уксусный альдегид. Значит, ацетальдегид блокируется и исключается из последующего процесса:
СН3СНО + NaHSO3 → СН3СНОНSO3Na (3.11)
В таких условиях акцептором водорода, оторванного от НАДН2, является фосфоглицериновый альдегид, который превращается в глицерин-3-фосфат, а затем дефосфорилируется с образованием глицерина:
С6Н12О6 → С3Н5(ОН)3 + СН3СНОНSO3Na + СО2 (3.12)
Сходный вариант спиртового брожения происходит при подщелачивании среды:
2С6Н12О6 + Н2О → 2 С3Н5(ОН)3 + СН3СН2ОН + СН3СООН + 2СО2 (3.13)
Маслянокислое брожение. Маслянокислое брожение – это процесс превращения бактериями в анаэробных условиях углеводов, спиртов и других органических веществ в масляную кислоту (бутират). Суммарное уравнение процесса:
С6Н12О6 → СН3СН2СН2СООН + 2 СО2 + Н2 + Эн. (3.14)
Маслянокислое брожение широко распространено в природе. Маслянокислые бактерии обитают там, где много органического вещества и нет доступа воздуха – в иловых отложениях водоемов, в почве, в скоплениях разлагающихся отбросов, в навозе, в сточных жидкостях и т.п. Развитее этих бактерий в почве, где достаточно воздуха, становится возможным благодаря симбиозу с аэробными бактериями, использующими кислород.
Маслянокислое брожение играет важную роль в цепи превращений органических веществ при минерализации.
Возбудители маслянокислого брожения относятся к роду Clostridium (C.butyricum, C.saccharobutyricum, C.amylobacter). Это строгие анаэробы. Они образуют споры, отличающиеся высокой устойчивостью. По отношению к источникам азота маслянокислые бактерии весьма неприхотливы. Они усваивают белковый, аминокислотный и аммонийный азот, а некоторые – азот воздуха.
Бактерии C.butyricum – возбудители типичного маслянокислого брожения. Это подвижные крупные грамположительные анаэробные палочки, споры располагаются центрально или субтерминально. Характерным является накопление гранулезы (специфического запасного углеводного вещества) перед образованием спор. Споры могут выдерживать кипячение в течение нескольких минут. Оптимальная температура развития бактерий 30…40оС, оптимальная рН - 6,9…7,4, при рН 4,5 они прекращают развитие.
Бактерии C.pasteurianumпо многим свойствам сходны с C.butyricum, но более кислотоустойчивые и не сбраживают крахмал. Они способны усваивать атмосферный азот.
Дата добавления: 2017-11-21; просмотров: 2602;
Поиск по сайту
Узнать еще
- I.1.6 ОСНОВНОЕ УРАВНЕНИЕ КИНЕТИЧЕСКОЙ ТЕОРИИ ГАЗОВ. ЗАКОН РАВНОМЕРНОГО РАСПРЕДЕЛЕНИЯ ЭНЕРГИИ ПО СТЕПЕНЯМ СВОБОДЫ
- I.3.3 МЕХАНИЧЕСКАЯ ЭНЕРГИЯ. ЗАКОН СОХРАНЕНИЯ МЕХАНИЧЕСКОЙ ЭНЕРГИИ
- II. Судовождение с использованием лоцманского метода и графического счисления пути судна.
- А) Спектр света и значение разного типа излучений
- А) Строительство гнезд (или использование других убежищ для икры).
- Автоматизированная информационная система организации перевозок грузов по безбумажной технологии с использованием электронной накладной (АИС ЭДВ)
- Автоматизированный асинхронный электропривод с использованием синхронных электромашинных преобразователей частоты.
- Автоматизированный электропривод с использованием ПЧ с ШИР.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
