ФИЗИОЛОГИЯ ПОЧЕК И МОЧЕВЫВОДЯЩИХ ПУТЕЙ
Почка- паренхиматозный, наиболее сложно устроенный орган в мочевой системе. Структурно-функциональными единицами ее являются нефроны, которые, в процессе образования мочи, обеспечивают все основные функции органа. К ним относятся: регуляция водно-электролитного баланса организма; задержка жизненно важных веществ, таких как белок и глюкоза; поддержание кислотно-основного баланса; экскреция продуктов обмена, водорастворимых токсинов, лекарств; регуляция осмотического и кровяного давления, эритро-поэза; эндокринная функция.
Регуляция водно-электролитного баланса организма. Почки позволяют человеку есть и пить в соответствии с его привычками без изменения состава жидкостных и электролитных параметров организма.
Кровоснабжение почек в норме составляет 20 % от сердечного выброса. Примерно 99 % почечного кровотока приходится на корковый и 1 % - на мозговой слой почек. Большинство нефронов расположено в корковом, наружном слое органа. Мозговой, внутренний слой почки содержит специализированные нефроны в юкстамедуллярной области, лежащей на границе мозгового слоя. Эти нефроны обладают высокой концентрационной способностью, механизм которой будет рассмотрен ниже.
Почка - поистине уникальный орган, имеющий два капиллярных бассейна, состоящих из двух типов капилляров: клубочковых, находящихся под высоким
давлением и осуществляющих фильтрацию, и околоканальцевых (паратубуляр-ных), с низким давлением. Все это позволяет фильтровать и реабсорбировать большие объемы жидкости.
Нефрон- структурно-функциональная единица почки. Каждая почка содержит около миллиона нефронов. Нефрон состоит из клубочка и канальцев (рис. 2.6). Канальцы разделены на следующие отделы: проксимальный каналец, петля мозгового слоя (петля Генле) и дистальный каналец, впадающий в собирательную трубку. Моча образуется в результате трехфазного процесса: 1) простой фильтрации; 2) избирательной реабсорбции; 3) пассивной реабсорбции и экскреции.
Фильтрация происходит через полупроницаемую стенку капилляров клубочка, которая в основном непроницаема для белков и крупных молекул. Таким образом, фильтрат не содержит белка и клеточных элементов. Клубоч-ковый фильтрат формируется путем продавливания крови через капилляры клубочков. Движущей силой фильтрации является гидростатическое давление, которое регулируется приносящей и выносящей артериолами и обеспечивается артериальным давлением. Каждую минуту фильтруется около 20 % почечного
плазмотока (125 мл/мин), что равняется скорости клубочковой фильтрации.
Для сохранения относительно постоянных величин почечного кровотока и скорости клубочковой фильтрации в клубочках поддерживается достаточно постоянное гидростатическое давление. При изменении артериального давления происходит сокращение или расширение афферентной и эфферентной артериол - сосудов мышечного типа, входящих и выходящих из каждого клубочка. Этот процесс называется ауторе-гуляцией.
Ауторегуляция скорости клубочко-вой фильтрации достигается посредством саморегуляции почечного кровотока и механизма обратной связи, известного как клубочково-канальцевое равновесие.
Клубочково-канальцевое равновесие. При уменьшении скорости клубочко-вой фильтрации происходит снижение тока жидкости в канальцах и увеличивается время реабсорбции ионов натрия и хлора. Уменьшение количества ионов натрия и хлора, достигающих дисталь-ного канальца, ведет к снижению сопротивления афферентных артериол
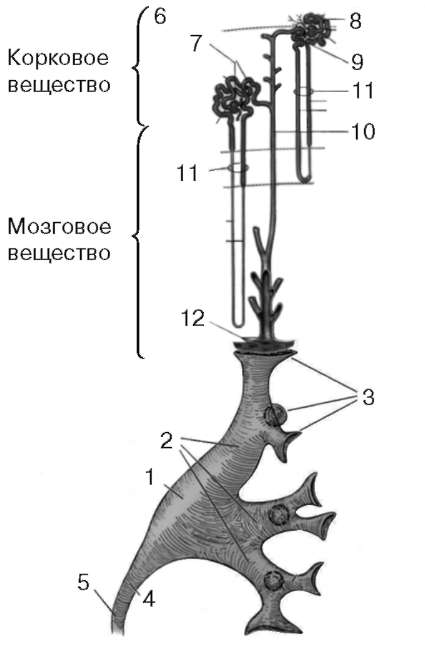
Рис. 2.6.Образование и выведение мочи:
1 - почечная лоханка; 2 - большие чашки; 3 - малые чашки; 4 - пиелоуретеральный сегмент; 5 - мочеточник; 6 - капсула; 7 - тельце почки; 8 - проксимальный извитой каналец; 9 - дистальный извитой каналец; 10 - собирательная трубочка;
11 - петля нефрона; 12 - решетчатое поле почечного сосочка
и сопровождается повышением почечного кровотока. При этом усиливается секреция ренина из юкстагломерулярного аппарата, что стимулирует выброс ангиотензина II, вызывающего сокращение эфферентных артериол. Повышение гидростатического давления в капиллярах клубочков возвращает скорость клубочковой фильтрации к ее нормальным значениям.
Юкстагломерулярный комплекс состоит из клеток плотного пятна (macula densa) - юкстагломерулярных клеток, являющихся специализированным эпителием дистального канальца, чувствительного к концентрации ионов натрия и способного влиять на клетки гладкой мускулатуры стенок афферентной и эфферентной артериол. Клетки macula densa также секретируют ренин - фермент, конвертирующий сывороточный белок ангиотензиноген в ангиотензин I. Впоследствии ангиотензин-превращающий фермент, который образуется в небольших количествах в легких, проксимальных канальцах и других тканях, превращает ангиотензин I в ангиотензин II, вызывающий вазоконстрикцию и повышающий артериальное давление. Ангиотензин II также стимулирует кору надпочечников, повышая секрецию альдостерона, который в свою очередь вызывает задержку воды и натрия, увеличивая объем циркулирующей крови.
Приведенная схема поддержания клубочково-канальцевого равновесия представляет собой систему отрицательной обратной связи. Другими словами, начальным стимулом системы является падение объема циркулирующей крови, приводящее к снижению перфузионного давления почек. Когда объем циркулирующей крови, перфузия почек и скорость клубочковой фильтрации восстановились, система отвечает снижением или выключением ответа на исходный стимул.
Селективная, или пассивная, реабсорбция. Функция почечных канальцев - селективная реабсорбция 99 % клубочкового фильтрата. Проксимальный каналец абсорбирует 60 % всех растворенных веществ, в том числе 100 % глюкозы и аминокислот, 90 % бикарбоната и 80-90 % неорганического фосфора и воды.
Реабсорбция происходит посредством активного и пассивного транспорта. Активный транспорт требует энергии для перемещения веществ против электрохимического или концентрационного градиента. Это основная детерминанта потребления кислорода почками. Посредством пассивного транспорта происходит реабсорбция веществ по электрохимическому и концентрационному градиентам или по градиенту давлений.
В основном реабсорбция осуществляется посредством активного транспорта веществ и свободного перемещения воды по принципу осмоса. При активной реабсорбции веществ происходит снижение их концентраций и, следовательно, падение осмотической активности в просвете канальца. Затем из-за присутствия осмотических сил вода перемещается из канальца в интерстиций, где концентрация осмотически активных веществ выше.
Петля Генле- это часть канальца, погружающаяся, или «изгибающаяся», из коркового слоя в мозговой (нисходящее колено) и затем возвращающаяся в кору почек (восходящая колено). Именно в этой части канальца моча при необходимости концентрируется. Это возможно благодаря высокой концентрации веществ в интерстиции мозгового слоя, которая поддерживается за счет наличия «противоточно-поворотной системы». Противоточно-поворотная система
поддерживает высокий осмотический градиент интерстиция мозгового слоя, что позволяет почкам концентрировать мочу. Петля Генле - это противоточ-но-поворотный множитель, а vasa recta (отдел перикапиллярной системы, входящий в мозговой слой в области высокой концентрации абсорбированных из первичной мочи веществ) - это противоточно-поворотный обменник, механизм которого описан ниже.
Функции различных частей петли Генле:
Нисходящее колено петли Генле относительно непроницаемо для растворенных веществ и хорошо проницаемо для воды, перемещаемой из канальца по осмотическому градиенту: жидкость в канальце становится гиперосмолярной.
Тонкий сегмент восходящего колена петли Генле практически непроницаем для воды, но в то же время проницаем для растворенных веществ, особенно ионов натрия и хлора, которые перемещаются по концентрационному градиенту из просвета канальца, жидкость в котором вначале становится изотонич-ной, а затем гипотоничной по мере выхода из нее ионов. Мочевина, абсорбировавшаяся в интерстиций мозгового слоя почки из собирательной трубки, диффундирует в восходящее колено. Это поддерживает концентрацию мочевины в интерстиции мозгового слоя, играя важную роль в процессе концентрации мочи.
Толстый сегмент восходящего колена петли Генле и начальный отдел дис-тального канальца непроницаемы для воды. Однако здесь происходит активный транспорт ионов натрия и хлора из просвета канальца, вследствие чего жидкость этого отдела канальца становится крайне гипотоничной.
Дистальный каналец и собирательная трубка: конечная концентрация мочи зависит от количества антидиуретического гормона, секретируемого задней долей гипофиза. В присутствии антидиуретического гормона дистальный каналец и собирательная трубка становятся проницаемыми для воды. При прохождении собирательной трубки через мозговой слой с высокой интерстициаль-ной концентрацией веществ вода выходит из просвета трубки и формируется концентрированная моча. При отсутствии антидиуретического гормона стенки дистального канальца становятся непроницаемыми для воды; таким образом формируется большое количество разведенной мочи.
Имеется тесная связь между гипоталамусом и задним гипофизом. В гипоталамусе присутствуют клетки-осморецепторы, чувствительные к изменению осмотического давления крови. При высоком потреблении воды наблюдается снижение осмотического давления крови, а при ее дефиците, соответственно, идет обратный процесс. При повышении осмотического давления крови нервные импульсы из гипоталамуса стимулируют задний гипофиз и усиливают секрецию антидиуретического гормона. В результате выработки антидиуретического гормона снижается потеря воды почками, так как она реабсорбируется в собирательных трубках.
Vasa Recta взаимодействует с петлей Генле посредством сложного механизма, направленного на концентрацию мочи путем противоточно-поворотного обмена. При отсутствии vasa recta высокая концентрация веществ в мозговом слое вымывалась бы током крови. Вещества диффундируют из сосудов, несущих кровь прямо в корковый слой и в сосуды, спускающиеся в мозговой слой,
в то время как вода делает противоположное: движется из нисходящих сосудов в восходящие. Эта система посредством подобного шунта позволяет веществам и воде рециркулировать внутри мозгового слоя.
Поддержание кислотно-основного состояния. Легкие и почки в совокупности поддерживают рН крови и внеклеточной жидкости в пределах 7,35-7,45 (34- 46 нмоль/л - концентрация Н+). Углекислый газ (СО2), растворенный в крови, является кислотой и элиминируется легкими. Почки же удаляют связанную кислоту посредством трех процессов: канальцевой секреции кислоты, клубоч-ковой фильтрации буферов, связанных с Н+, и образования аммиака.
1. Канальцевая секреция кислоты: бикарбонат натрия фильтруется в клубочке, реабсорбируясь затем в проксимальном канальце. Натрий абсорбируется посредством Na+/Н+-ионной помпы, обменивая Na+ на Н+-ионы на мембранах эпителия проксимального канальца.Na+/К+-помпа продвигает натрий через клетку из первичной мочи в обмен на калий.
2. Клубочковая фильтрация буферов, связанных с Н+:
А. Основная часть фильтруемого бикарбоната реабсорбируется (90 % в проксимальном канальце). Н+, высвобожденный при канальцевой секреции кислоты (см. выше), соединяясь с бикарбонатом (HCO3), формирует углекислоту:
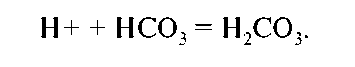
Карбоангидраза, присутствующая в клетках проксимального канальца, катализирует реакцию расщепления угольной кислоты на СО2 и Н2О. СО2 диффундирует в эпителиальную клетку и в присутствии карбоангидразы образует углекислоту. Последняя ионизируется до Н+ и НСО3. Н+ затем выкачивается из клетки в просвет канальца Na+/Н+-помпой, а натрий возвращается в плазму Na+/К+-помпой (см. выше); вода же абсорбируется пассивно.
B. Другие буферы, включая неорганический фосфат (НРО3), ураты и ионы креатинина, в дистальном отделе нефрона экскретируются в мочу как кислоты, если они связаны с Н+.
3. Аммиак (NH3) образуется ферментативно из глутамина и других аминокислот и секретируется в канальцы нефрона. Аммиак в сочетании с секретирован-ным в мочу Н+-ионом формирует недиффундирующий аммиак-ион (NH4-), выводимый с мочой.
Экскреция продуктов метаболизма. Фильтрация их происходит во время продвижения крови по клубочку. Некоторые ненужные организму вещества и чужеродные субстанции, например лекарственные препараты, не могут быть выведены из организма путем фильтрации. Такие вещества секретируются в канальцы нефрона и выделяются из организма с мочой.
Гормоны и почки. Ренин повышает продукцию ангиотензина II, высвобождаемого при снижении внутрисосудистого объема, например при кровопотере или дегидратации. Это ведет к:
■ констрикции эфферентных артериол для поддержания скорости клубоч-ковой фильтрации за счет повышения фильтрационного давления в клубочке;
■ выбросу альдостерона из коркового вещества надпочечников;
■ повышению секреции антидиуретического гормона задней долей гипофиза;
■ положительному инотропному действию на сердце и артериальной вазо-констрикции.
Альдостерон усиливает реабсорбцию ионов натрия и воды в дистальном канальце и собирательной трубочке, где Na+ обменивается на К+ и ионы водорода специфическими клеточными помпами. Секреция альдостерона усиливается при снижении концентрации Na+ в сыворотке крови. Это может произойти, например, при потере большого объема желудочного сока, так как желудочный сок содержит значительное число ионов натрия, хлора, водорода и калия. Следовательно, невозможно корригировать возникающий алкалоз и гипокалиемию без предварительного возмещения ионов натрия физиологическим раствором.
Предсердный натрийуретический пептид секретируется при повышении давления в предсердиях, например при сердечной недостаточности или жидкостной перегрузке. Предсердный натрийуретический пептид приводит к повышению потерь натрия, хлоридов и воды преимущественно за счет повышения скорости клубочковой фильтрации.
Антидиуретический гормон повышает проницаемость стенок дистального канальца и собирательной трубки для воды и таким образом концентрирует мочу. С другой стороны, при сниженной секреции антидиуретического гормона формируется значительное количество «разведенной» мочи. Подобная ситуация возникает преимущественно при падении концентрации натрия в плазме крови после потребления больших объемов воды. Снижение уровня натрия контролируется осморецепторами. При кровопотере или дегидратации гормоны взаимодействуют между собой, что играет роль в поддержании нормального внутрисосудистого объема.
К другим веществам, синтезируемым почками, относятся 1,25-дигидрок-си-витамин D (наиболее активная форма витамина D), обеспечивающий абсорбцию кальция из кишечника, и эритропоэтин, стимулирующий продукцию эритроцитов. Продукция этих веществ снижается при почечной недостаточности.
Физиология мочевыделения. Образующаяся в почечных канальцах моча выделяется в почечную чашечку, а затем в фазе ее систолы попадает в почечную лоханку. Последняя постепенно заполняется мочой, и по достижении порога раздражения возникают импульсы от барорецепторов, сокращается мускулатура почечной лоханки, раскрывается просвет мочеточника и моча благодаря сокращениям его стенки продвигается в мочевой пузырь.
Функция нижних мочевыводящих путей состоит из двух фаз - накопления мочи (резервуарная функция) и ее эвакуации. При этом отмечается определенный, но равномерный режим работы мочевого пузыря и запирательного аппарата, то есть чередование между его заполнением и опорожнением. При физиологических условиях заполнение мочевого пузыря происходит подсознательно и очень медленно (у взрослого человека в течение 2-4 часов), а эвакуация осуществляется по желанию и заканчивается в течение 20-30 секунд. У ребенка до 1,5-2 лет мочеиспускание рефлекторное. Во время роста, с развитием проводящих путей между центрами спинного мозга и головным мозгом, мочеиспускание становится контролируемым.
Процессы накопления и эвакуации мочи обеспечиваются определенными анатомическими образованиями и составляют единую функциональную систему. Удержание мочи обеспечивают следующие образования: шейка мочевого пузыря и проксимальная уретра, часто рассматриваемая как внутренний сфинктер (или гладкомышечный сфинктер мочевого пузыря), наружный сфинктер (или поперечно-полосатый сфинктер уретры), мышцы тазового дна. Накопление мочи обеспечивается мышцами мочевого пузыря. Функционально процесс удержания мочи связан со сложным комплексом физиологических механизмов как запирательного аппарата, так и мочевого пузыря, что обеспечивается рефлекторными актами и чисто механическими компонентами.
Для лучшего понимания нормальной функции и дисфункции нижних мочевых путей необходимо изложить особенности их нервной регуляции. Афферентная иннервация мочевого пузыря и уретры осуществляется за счет рецепторов, чувствительных к болевому, температурному и прессорному воздействию. Чувствительные рецепторы имеются во всех слоях мочевого пузыря, но наибольшее их количество расположено в области треугольника Льето. Выделяются резко специализированные рецепторы мочевого пузыря, реагирующие на быстрые изменения его объема, и окончания, воспринимающие медленные изменения внутреннего давления. По степени адаптации к наполнению различают фазные и тонические рецепторы мочевого пузыря. Важную роль в акте мочеиспускания играют рецепторы, заложенные в стенке уретры, особенно в ее проксимальном отделе, а также чувствительные рецепторы поперечно-полосатых мышц уретры и промежности.
Все нервные импульсы, генерируемые в нижних мочевыводящих путях, поступают в центральные отделы нервной системы, за счет чего осуществляется координированный акт мочеиспускания. К нервным центрам относятся:
■ интрамедиолатеральные клетки стволов и клетки вентральных рогов серого вещества сакрального отдела спинного мозга;
■ ретикулярная формация ствола мозга;
■ мозжечок, который получает нервные импульсы от детрузора и мышц тазового дна через спиноцеребеллярный тракт;
■ передняя группа ядер гипоталамуса;
■ базальные ганглии, образования стволовой клетки мозга: хвостатое ядро, чечевицеобразное ядро, черная субстанция, красное ядро;
■ зрительные бугры (неспецифические таламические ядра, которые расположены во внутреннем слое);
■ кора головного мозга - является центром координации акта мочеиспускания. Эфферентная система регуляции акта мочеиспускания начинается от коры
головного мозга, причем корковые центры регуляции акта мочеиспускания - понятие функционально-динамическое, имеющее, помимо постоянного анатомического представления, многочисленные условно-рефлекторные связи. Из клеток 5-го слоя коры больших полушарий возникают нисходящие волокна, идущие к субкортикальным образованиям и дальше к спинному мозгу. Однако считается, что нет убедительных доказательств наличия прямого кортикоспи-нального пути, а существуют многочисленные короткие пути и промежуточные пункты.
В медиальной ретикулярной формации начинаются аксоны, проходящие через вентральный ретикулоспинальный путь к спинным центрам мочеиспускания. Спинномозговые центры мочеиспускания располагаются в тора-колюмбальном и сакральном отделах спинного мозга, что соответствует парасимпатическим и симпатическим участкам вегетативной нервной системы. Моторные преганглионарные симпатические нейроны начинаются в клетках, которые локализуются в интермедиолатеральных ядрах сегментов от Th 12 до L 2, и участвуют в формировании чревных нервов и ганглиев чревного сплетения. Преганглионарные волокна проходят через ганглии паравертебрального ствола и в составе чревного нерва заканчиваются в узлах пузырных сплетений. Постганглионарные волокна направляются к детрузору, шейке мочевого пузыря и треугольнику Льето. Адренергические рецепторы размещаются в нижних мочевыводящих путях неравномерно: α-адренорецепторы преобладают в области шейки мочевого пузыря, проксимальной уретры, предстательной железе; Р-адренергические рецепторы располагаются в области тела мочевого пузыря. Рецепторы высвобождают адренергические нейромедиаторы (норад-реналин и адреналин). В мочевой системе преобладаютα1А- и α1D -адрено-рецепторы. На рис. 2.7 представлена локализация адренорецепторов в мочевом пузыре, простате и уретре.
Парасимпатические эфферентные группы, идущие к нижним мочевым путям, начинаются клеточными телами в сакральных парасимпатических ядрах (интермедиалатеральном сером веществе) от S2-S4 сегментов. Моторные волокна через вентральный корешок и далее в составе тазовых и подчревных нервов направляются к пузырному сплетению. Холинергиче-ские рецепторы преобладают в области тела мочевого пузыря и почти не встречаются в проксимальной уретре.
Возбуждение парасимпатической нервной системы ведет к сокращению детрузора. Возбуждение симпатической нервной системы вызывает неоднозначные функциональные изменения. Так, моторный β-адренер-гический эффект вызывает расслабление детрузора, раскрытие устьев мочеточников; α -адренергическое влияние вызывает сокращение три-гональной мышцы и интрамуральной мускулатуры мочеточников, повышает тонус области внутреннего сфинктера и проксимальной уретры.
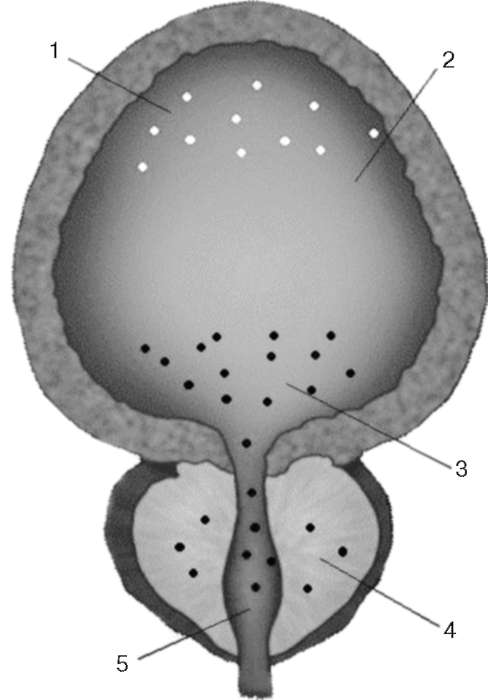
Рис. 2.7.Локализация адренорецепторов в мочевом пузыре, простате и уретре: 1 - β-адренорецепторы; 2 - мочевой пузырь; 3 -α -адренорецепторы; 4 - предстательная железа; 5 - проксимальная уретра
Таким образом, суммарное действие симпатической иннервации на нижние мочевыводящие пути заключается в постоянном поддержании тонуса внутреннего сфинктера и проксимальной уретры, раскрытии устьев мочеточников, расслаблении детрузора по мере наполнения мочевого пузыря. Прекращение симпатического влияния совпадает с активацией рефлекса мочеиспускания.
Помимо симпатической и парасимпатической иннервации акт мочеиспускания регулирует и соматическая нервная система. Эфферентные нейроны, идущие от серого вещества переднего рога S2-S4 сегментов спинного мозга через срамное сплетение и срамной нерв, заканчиваются в области поперечнополосатого наружного сфинктера и в мускулатуре тазового дна.
Акт мочеиспускания обеспечивается за счет сложной рефлекторной регуляции центральной и периферической нервной системы. Рефлекторные механизмы скоординированы во времени. Активность рефлекса отражает сумму всех возбуждающих и тормозящих нервных импульсов, воздействующих на аппарат нижних мочевыводящих путей.
Акт мочеиспускания осуществляется следующим образом. Сокращающийся m. detrusor urinae выжимает мочу из мочевого пузыря в мочеиспускательный канал, открывающийся благодаря расслаблению своих сфинктеров: непроизвольного (m. sphincter vesicae) и произвольного (m. sphincter urethrae). При этом у мужчин происходит также расслабление мышечной части предстательной железы, выполняющей функции третьего (непроизвольного) сфинктера. Закрытие мочевого пузыря происходит при расслаблении m. detrusor и сокращении названных сфинктеров.
Возбуждение центра мочеиспускания вызывает импульсацию в парасимпатических волокнах тазовых внутренностных нервов (nn. splanchnici pelvici), а мышца наружного сфинктера иннервируется соматическим нервом - ветвью полового нерва (n. pudendus).
Движение мочи по мочеиспускательному каналу играет важную роль в акте мочеиспускания: оно рефлекторно по афферентным волокнам полового нерва стимулирует сокращение мочевого пузыря. Поступление мочи в задние отделы мочеиспускательного канала и его растяжение способствуют сокращению мышцы мочевого пузыря. Передача афферентных и эфферентных импульсов этого рефлекса осуществляется по подчревному нерву (n. hypogastricus).
2.4. ФИЗИОЛОГИЯ МУЖСКИХ ПОЛОВЫХ ОРГАНОВ
Дата добавления: 2021-06-28; просмотров: 614;
Поиск по сайту
Узнать еще
- Алгоритм ОКАЗАНИЯ ПЕРВОЙ ПОМОЩИ при обструкции дыхательных путей инородным телом
- Анализ путей и стратегий станкостроения
- Анализ результатов обследования и выбор путей коррекционного воздействия
- АНАТОМИЯ И ФИЗИОЛОГИЯ ОРГАНОВ ДЫХАНИЯ.
- Анатомия и физиология поджелудочной железы
- Анатомия и физиология почек
- Анатомия и физиология сердца
- АНАТОМИЯ И ФИЗИОЛОГИЯ ЧЕЛОВЕКА КАК НАУКА

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
