Выделяют две основные функции спинного мозга: проводниковую и рефлекторную.
Проводниковая функция обеспечивает связь нейронов спинного мозга друг с другом или с вышележащими отделами ЦНС.
Рефлекторная функция позволяет реализовать все двигательные рефлексы тела, рефлексы внутренних органов, мочеполовой системы, терморегуляции и т.д. Собственная рефлекторная деятельность спинного мозга осуществляется сегментарными рефлекторными дугами.
Введем некоторые важные определения. Минимальная сила раздражения, которая вызывает рефлекс, называется порогом (или пороговым стимулом) данного рефлекса. Всякий рефлекс имеет рецептивное поле, т. е. совокупность рецепторов, раздражение которых вызывает рефлекс с наименьшим порогом.
При изучении движений приходится расчленять сложный рефлекторный акт на отдельные относительно простые рефлексы. Вместе с тем следует помнить, что в естественных условиях отдельный рефлекс выступает только как элемент сложной деятельности.
Спинальные рефлексы делят:
Во-первых, по рецепторам, раздражение которых вызывает рефлекс:
а) проприоцептивные (собственные) рефлексы с самой мышцы и связанных с ней образований. Они имеют простейшую рефлекторную дугу. Рефлексы, возникающие с проприорецептров, участвуют в формировании акта ходьбы и регуляции мышечного тонуса.
б) висцероцептивные рефлексы возникают с рецепторов внутренних органов и проявляются в сокращении мышц брюшной стенки, грудной клетки и разгибателей спины. Возникновение висцеромоторных рефлексов связано с конвергенцией висцеральных и соматических нервных волокон к одним и тем же интернейронам спинного мозга,
в) кожные рефлексы возникают при раздражении рецепторов кожи сигналами внешней среды.
Во-вторых, по органам:
а) рефлексы конечностей;
б) брюшные рефлексы;
в) яичковый рефлекс;
г) анальный рефлекс.
Простейшими спинальными рефлексами, которые можно легко наблюдать, являются сгибательный и разгибательный. Под сгибанием следует понимать уменьшение угла данного сустава, а под разгибанием его увеличение. Сгибательные рефлексы широко представлены в движениях человека. Характерным для этих рефлексов является большая сила, которую они могут развивать. Вместе с тем они быстро утомляются. Разгибательные рефлексы также широко представлены в движениях человека. Например, к ним относятся рефлексы поддержания вертикальной позы. Эти рефлексы в отличие от сгибательных значительно более устойчивы к утомлению. Действительно, мы можем долго ходить и стоять, но для выполнения длительной работы, например поднимания рукой гири, наши физические возможности значительно более ограничены.
Универсальный принцип рефлекторной деятельности спинного мозга получил название общего конечного пути. Дело в том, что соотношение количества волокон в афферентных (задних корешках) и эфферентных (передних корешках) путях спинного мозга составляет примерно 5:1. Ч. Шеррингтон образно сравнивал этот принцип с воронкой, широкую часть которой составляют афферентные пути задних корешков, а узкую эфферентные пути передних корешков спинного мозга. Нередко территория конечного пути одного рефлекса перекрывается с территорией конечного пути другого рефлекса. Другими словами, разные рефлексы могут соревноваться за занятие конечного пути. Это можно проиллюстрировать таким примером. Представим себе, что собака убегает от опасности и ее в это время кусает блоха. В данном примере за общий конечный путь - мышцы задней лапы - соревнуются два рефлекса: один-чесательный, а другой - рефлекс ходьбы-бега. В одни моменты может пересиливать чесательный рефлекс, и собака останавливается и начинает чесаться, но затем вновь верх может взять рефлекс ходьбы-бега, и собака возобновит бег.
Как уже указывалось, при осуществлении рефлекторной деятельности отдельные рефлексы взаимодействуют друг с другом, образуя функциональные системы. Один из самых важных элементов функциональной системы - обратная афферентация, благодаря которой нервные центры как бы оценивают, как выполняется реакция, и могут внести в нее необходимые коррективы.
Рассмотрим рефлексы конечностей.
Рефлексы на растяжение мышцы. Известны два вида рефлекса на растяжение: фазный (быстрый) и тонический (медленный). Примером фазного рефлекса может служить коленный рефлекс, который возникает при легком ударе по сухожилию мышцы в подколенной чашечке. Рефлекс на растяжение предотвращает чрезмерное растяжение мышцы, которая как бы сопротивляется растягиванию. Этот рефлекс возникает как ответная реакция мышцы на стимуляцию ее рецепторов, поэтому его часто обозначают как собственный рефлекс мышцы. Быстрое растяжение мышцы, всего на несколько миллиметров механическим ударом по ее сухожилию приводит к сокращению всей мышцы и разгибанию голени.
Путь этого рефлекса следующий:
- мышечные рецепторы четырехглавой мышцы бедра;
- спинальный ганглий;
- задние корешки;
- задние рога III поясничного сегмента;
- мотонейроны передних рогов того же сегмента;
- волокна четырехглавой мышцы бедра.
Реализация этого рефлекса была бы невозможна, если бы одновременно с сокращением мышц-разгибателей не расслаблялись мышцы-сгибатели. Поэтому во время разгибательного рефлекса мотонейроны мышц-сгибателей тормозятся с помощью вставочных тормозных клеток Реншоу (реципрокное торможение). Фазные рефлексы участвуют в формировании ходьбы. Рефлекс на растяжение свойствен всем мышцам, но у мышц-разгибателей, они хорошо выражены и легко вызываются.
К фазным рефлексам на растяжение относят также ахиллов рефлекс, вызываемый легким ударом по ахиллову сухожилию, и локтевой рефлекс вызываемый ударом молоточка по сухожилию четырехглавой мышцы.
Тонические рефлексы возникают при длительном растяжении мышц, их главное назначение – поддержание позы. В положении стоя тоническое сокращение мышц-разгибателей предотвращает сгибание нижних конечностей под действием сил гравитации и обеспечивает сохранение вертикального положения. Тоническое сокращение мышц спины обеспечивает осанку человека. Тоническое сокращение скелетных мышц является фоновым для осуществления всех двигательных актов, осуществляемых с помощью фазных сокращений мышц. Примером тонического рефлекса на растяжение может быть собственный рефлекс икроножной мышцы. Это одна из главных мышц, благодаря которой поддерживается вертикальная поза человека.
Более сложно организованы рефлекторные ответы, выражающиеся в координированном сгибании и разгибании мышц конечностей. Примером являются сгибательные рефлексы, направленные на избежание различных повреждающих воздействий. Рецептивное поле сгибательного рефлекса достаточно сложно и включает различные рецепторные образования и различные по скорости проведения афферентные пути. Сгибательный рефлекс возникает при раздражении болевых рецепторов кожи, мышц и внутренних органов. Вовлекаемые при этих раздражениях афферентные волокна имеют широкий спектр скоростей проведения - от миелинизированных волокон группы А до немиелинизированных волокон группы С. Все разнообразные афферентные волокна, импульсация по которым приводит к развитию сгибательного рефлекса, объединяют под названием афферентов сгибательного рефлекса.
Сгибательные рефлексы отличаются от собственных рефлексов мышц не только большим числом синаптических переключений на пути к мотонейронам, но и вовлечением ряда мышц, координированное сокращение которых обусловливает движение целой конечности. Одновременно с возбуждением мотонейронов, иннервирующих мышцы-сгибатели, происходит реципрокное торможение мотонейронов мышц-разгибателей.
При достаточно интенсивном раздражении рецепторов нижней конечности происходят иррадиация возбуждения и вовлечение в реакцию мышц верхней конечности и туловища. При активации мотонейронов противоположной стороны тела наблюдается не сгибание, а разгибание мышц противоположной конечности - перекрестный разгибательный рефлекс.
Рис. Схема сгибательного рефлекса.
(Фиолетовым закрашены тормозные интернейроны)
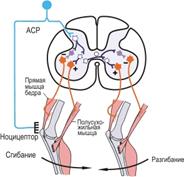
Еще более сложный характер имеют рефлексы позы – перераспределение мышечного тонуса, возникающее при изменении положения тела или отдельных его частей. Они представляют собой большую группу рефлексов. Сгибательный тонический рефлекс позы можно наблюдать у лягушки и у млекопитающих, для которых характерно подогнутое положение конечностей (кролик).
Для большинства млекопитающих и человека главное значение для поддержания положения тела играет не сгибательный, а разгибательный рефлекторный тонус. На уровне спинного мозга особенно важную роль в рефлекторной регуляции разгибательного тонуса играют шейные позные- рефлексы. Их рецепторы содержатся в мышцах шеи. Рефлекторная дуга полисинаптическая, замыкается на уровне I-III шейных сегментов. Импульсы от этих сегментов передаются мышцам туловища и конечностей, вызывая перераспределение их тонуса. Имеется две группы этих рефлексов – возникающие при наклоне и при повороте головы.
Первая группа шейных позных рефлексов существует только у животных и возникает при наклоне головы вниз (кпереди). При этом увеличивается тонус мышц-сгибателей передних конечностей и тонус мышц-разгибателей задних конечностей, в результате чего передние конечности сгибаются, а задние разгибаются. При наклоне головы вверх (кзади) возникают противоположные реакции – передние конечности разгибаются вследствие увеличения тонуса их мышц-разгибателей, а задние конечности сгибаются вследствие повышения тонуса их мышц-сгибателей. Эти рефлексы возникают с проприорецепторов мышц шеи и фасций, покрывающих шейный отдел позвоночника. В условиях естественного поведения они увеличивают животному шанс достать пищу, находящуюся выше или ниже уровня головы.
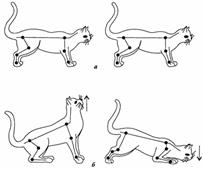
Рис. Позные шейные рефлексы у кошки с удаленным вестибулярным аппаратом. A - до изменения положения головы; б- при пассивных поднимании (↑) и опускании (↓) головы.
Рефлексы позы верхних конечностей у человека утрачены. Рефлексы нижних конечностей выражаются не в сгибании или разгибании, а в перераспределении мышечного тонуса, обеспечивающего сохранение естественной позы.
Вторая группа шейных позных рефлексов возникает с тех же рецепторов, но только при поворотах головы вправо или влево. При этом повышается тонус мышц-разгибателей обеих конечностей на стороне, куда повернута голова, и повышается тонус мышц-сгибателей на противоположной стороне. Рефлекс направлен на сохранение позы, которая может быть нарушена вследствие изменения положения центра тяжести после поворота головы. Центр тяжести смещается в сторону поворота головы – именно на этой стороне повышается тонус мышц-разгибателей обеих конечностей. Подобные рефлексы наблюдаются и у человека.
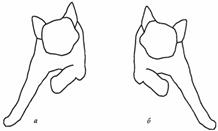
Рис. Изменение тонуса мышц конечностей при наклоне головы вправо (а) и влево (б).
На уровне спинного мозга замыкаются также ритмические рефлексы – многократное повторное сгибание и разгибание конечностей. Примерами могут служить чесательный и шагательный рефлексы. Ритмические рефлексы характеризуются координированной работой мышц конечностей и туловища, правильным чередованием сгибания и разгибания конечностей наряду с тоническим сокращением приводящих мышц, устанавливающих конечность в определенное положение к кожной поверхности.
Брюшные рефлексы (верхний, средний и нижний) проявляются при штриховом раздражении кожи живота. Выражаются в сокращении соответствующих участков мускулатуры стенки живота. Это защитные рефлексы. Для вызова верхнего брюшного рефлекса раздражение наносят параллельно нижним ребрам непосредственно под ними, дуга рефлекса замыкается на уровне VIII-IX грудного сегмента спинного мозга. Средний брюшной рефлекс вызывают раздражением на уровне пупка (горизонтально), дуга рефлекса замыкается на уровне IX-X грудного сегмента. Для получения нижнего брюшного рефлекса раздражение наносят параллельно паховой складке (рядом с ней), дуга рефлекса замыкается на уровне XI-XII грудного сегмента.
Кремастерный (яичковый) рефлекс заключается в сокращении m. сremaster и поднимании мошонки в ответ на штриховое раздражение верхней внутренней поверхности кожи бедра (кожный рефлекс), это также защитный рефлекс. Его дуга замыкается на уровне I-II поясничного сегмента.
Анальный рефлекс выражается в сокращении наружного сфинктера прямой кишки в ответ на штриховое раздражение или укол кожи вблизи заднего прохода, дуга рефлекса замыкается на уровне IV-V крестцового сегмента.
Вегетативные рефлексы. Кроме рассмотренных выше рефлексов, которые относятся к категории соматических, так как выражаются в активации скелетных мышц, спинной мозг играет важную роль в рефлекторной регуляции внутренних органов, являясь центром многих висцеральных рефлексов. Эти рефлексы осуществляются при участии нейронов вегетативной нервной системы, расположенных в боковых рогах серого вещества. Аксоны этих нервных клеток покидают спинной мозг через передние корешки и заканчиваются на клетках симпатических или парасимпатических вегетативных ганглиев. Ганглионарные нейроны в свою очередь посылают аксоны к клеткам различных внутренних органов, включая гладкие мышцы кишечника, сосудов, мочевого пузыря, к железистым клеткам, сердечной мышце. Вегетативные рефлексы спинного мозга осуществляются в ответ на раздражение внутренних органов и заканчиваются сокращением гладкой мускулатуры этих органов.
Нарушения функций спинного мозга.
В случаях раздражения и поражения задних корешков спинного мозга наблюдаются «стреляющие», опоясывающие боли на уровне метамера пораженного сегмента, снижение чувствительности всех видов, утрата или снижение рефлексов, вызываемых с метамера тела, который передает информацию в пораженный корешок.
В случаях изолированного поражения заднего рога утрачивается болевая и температурная чувствительность на стороне повреждения, а тактильная и проприоцептивная сохраняется, так как из заднего корешка аксоны температурной и болевой чувствительности идут в задний рог, а аксоны тактильной и проприоцептивной — прямо в задний столб и по проводящим путям поднимаются вверх. Вследствие того, что аксоны вторых нейронов болевой и температурной чувствительности идут на противоположную сторону через переднюю серую спайку спинного мозга, при повреждении этой спайки на теле симметрично утрачивается болевая и температурная чувствительность.
Поражение переднего рога и переднего корешка спинного мозга приводит к параличу мышц. Они теряют тонус, атрофируются, при этом исчезают рефлексы, связанные с пораженным сегментом.
В случае поражения боковых рогов спинного мозга исчезают кожные сосудистые рефлексы, нарушается потоотделение, наблюдаются трофические изменения кожи, ногтей. При одностороннем поражении парасимпатического отдела автономной нервной системы на уровне крестцовых отделов спинного мозга нарушений дефекации и мочеиспускания не наблюдается, так как корковая иннервация этих центров является двусторонней.
При травмах у человека в ряде случаев происходит полное или половинное пересечение спинного мозга.
При половинном латеральном повреждении спинного мозга развивается синдром Броун-Секара. Он проявляется в том, что на стороне поражения спинного мозга (ниже места поражения) развивается паралич двигательной системы вследствие повреждения пирамидных путей. На противоположной поражению стороне движения сохраняются. На стороне поражения (ниже места поражения) нарушается проприоцептивная чувствительность. Это обусловлено тем, что восходящие пути глубокой чувствительности идут по своей стороне спинного мозга до продолговатого мозга, где происходит их перекрест.
На противоположной стороне туловища (относительно повреждения спинного мозга) нарушается болевая чувствительность, так как проводящие пути болевой чувствительности кожи идут от спинального ганглия в задний рог спинного мозга, где переключаются на новый нейрон, аксон которого переходит на противоположную сторону. В итоге если повреждена левая половина спинного мозга, то исчезает болевая чувствительность правой половины туловища ниже повреждения.
Полную перерезку спинного мозга в экспериментах на животных производят для исследования влияния вышележащих отделов ЦНС на нижележащие. После полного пересечения спинного мозга возникает спинальный шок. Это явление заключается в том, что все центры ниже перерезки перестают организовывать присущие им рефлексы. Нарушение рефлекторной деятельности после пересечения спинного мозга у разных животных длится разное время. У лягушек оно исчисляется десятками секунд, у кролика рефлексы восстанавливаются через 10—15 мин, у собак отдельные рефлексы, например мышечного сокращения, восстанавливаются через несколько часов, другие — через несколько дней (рефлексы регуляции артериального давления), через недели восстанавливаются рефлексы мочеиспускания. У обезьян первые признаки восстановления рефлексов после перерезки спинного мозга появляются через несколько суток; у человека первые спинальные рефлексы восстанавливаются через несколько недель, а то и месяцев.
Следовательно, чем сложнее организация ЦНС, тем сильнее контроль вышележащих отделов мозга над нижележащими. То, что причиной шока является нарушение супраспинальных влияний, доказывается повторной перерезкой спинного мозга ниже места первой перерезки. В этом случае спинальный шок не возникает, рефлекторная деятельность спинного мозга сохраняется.
По истечении длительного периода времени после шока спинальные рефлексы резко усиливаются, что объясняется устранением тормозного влияния ретикулярной формации ствола мозга на рефлексы спинного мозга.
Дата добавления: 2017-01-16; просмотров: 3534;
Поиск по сайту
Узнать еще
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cущность организации и ее основные признаки
- I. Назначение унифицированных газозарядных станций и основные тактико-технические требования, предъявляемые к ним.
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I. ОСНОВНЫЕ ПОЛОЖЕНИЯ
- I. Политический режим: понятие, сущность и основные типы.
- I.2. Основные категории водопотребления промышленных предприятий и их особенности
- II. Митохондрии (строение и функции)

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
