Высшие жирные карбоновые кислоты (ВЖК), их классификация, номенклатура, свойства.
Многие известные высшие карбоновые кислоты были впервые выделены из жиров и в целом получили название жирных, а с учетом присущей им относительно длинной углеводородной цени их называют высшими жирными кислотами ( ВЖК).
В природе обнаружено свыше 500 представителей высших жирных кислот. Однако практически значимое распространение имеют немногим более 20 из них. В строении большинства природных жирных кислот наблюдаются определенные закономерности.
Как правило, природные высшие жирные кислоты характеризуются следующими признаками:
- являются монокарбоновыми;
- содержат четное число атомов углерода;
- имеют неразветвленный углеводородный радикал;
- имеют цис-конфигурацию двойных связей (если они содержатся). Однако существует и много исключений; например, известны кислоты с нечетным числом атомов углерода, с разветвленной углеводородной цепью, с транс-расположением двойных связей.
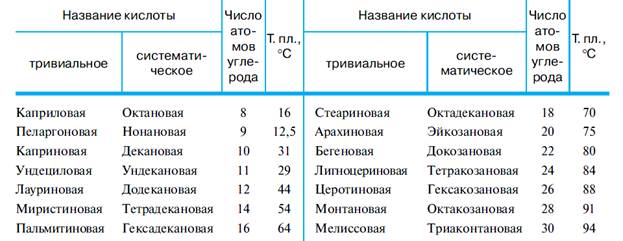
Строение и названия наиболее распространенных высших жирных кислот приведены в табл. 1 и 2.
Таблица 1. Насыщенные кислоты
Насыщенные жирные кислоты. Несмотря на большой диапазон возможной длины цепи, в составе липидов наиболее распространены кислоты с неразветвленной цепью, включающей 16, 18, 20 или 22 атома углерода (см. табл.1). Муравьиная, уксусная, пропановая — в липидах практически не встречаются; некоторые кислоты С4—С10 входят в состав липидов молока. Так называемые среднецепочечные жирные кислоты — лауриновая (С12) и миристиновая (С14) — характерны для липидов растений семейства пальм.
Наиболее распространенными являются пальмитиновая и стеариновая кислоты, особенно характерные для твердых жиров и некоторых твердых растительных масел (масло какао, пальмовое масло и др.). В жидких растительных маслах они содержатся в меньших количествах, при этом преобладающей является пальмитиновая кислота.
Примером насыщенной высшей жирной кислоты с разветвленной углеродной цепью является туберкулостеариновая (10-метилоктадекановая) кислота, впервые выделенная из туберкулезной палочки. В сливочном масле содержатся высшие жирные кислоты, имеющие в углеводородном радикале циклогексановое кольцо.
В углеводородном радикале насыщенных кислот все атомы углерода находятся в sp3-гибридизованном состоянии и имеют тетраэдрическую конфигурацию. За счет вращения вокруг сигма-связей С—С углеродная цепь может принимать различные конформации. Энергетически наиболее выгодной является зигзагообразная конформация, так как в этом случае атомы углерода цепи находятся в анти-бутановой (антиперипланарной) конформации, как это показано ранее на примере стеариновой кислоты.
Таблица 2. Ненасыщенные кислоты


Ненасыщенные жирные кислоты. Ненасыщенные высшие жирные кислоты распространены в природе более широко, чем насыщенные. Число атомов углерода в ненасыщенных жирных кислотах может быть различным, но преобладающими являются кислоты с 18 и 20 атомами углерода (см. табл. 2). В растительных маслах встречаются ненасыщенные кислоты с 22 атомами углерода (эруковая кислота в маслах из семян крестоцветных, например рапса).
Известны природные жирные кислоты с ненасыщенными пяти- и шестичленными циклами в углеводородных радикалах. Так, чаульмугровое масло, применяемое при лечении проказы, содержит в составе липидов высшие жирные кислоты с циклопентеновым кольцом — гиднокарповую и чаульмугровую (см. табл. 2).
В организме мононенасыщенные кислоты могут образовываться из насыщенных жирных кислот путем реакции дегидрирования. Основными синтезируемыми в организме ненасыщенными кислотами являются олеиновая и пальмитолеиновая (см. табл.2).
К полиеновым относят ненасыщенные высшие жирные кислоты, содержащие от двух до шести двойных связей. Каждая из них отделена от другой метиленовой группой. Отсюда и их название — метиленразделенные кислоты.
В молекулах ненасыщенных кислот, наряду с sp3-гибридизованными, содержатся sp2-гибридизованные атомы углерода. Поэтому участки углеводородной цепи относительно двойной связи могут иметь цис- или транс-расположение:
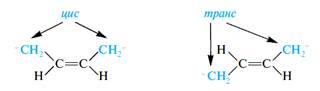
В ненасыщенных высших жирных кислотах природного происхождения двойные связи преимущественно имеют цис-конфигурацию, хотя это обычно менее выгодно, чем транс-расположение. Однако в данном случае цис-форма приводит к возможности формирования нужной вторичной структуры в липидной части клеточных мембран.
Ненасыщенные жирные кислоты могут подвергаться различным видам изомеризации, в частности пространственной — с изменением конфигурации двойных связей и позиционной — со смещением положения двойных связей.
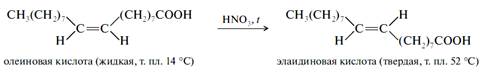
Пространственная (цис-транс) изомеризация легко происходит при нагревании, особенно в присутствии инициаторов радикальных реакций. Классическим примером такой изомеризации является получение элаидиновой кислоты при обработке олеиновой оксидами азота, образующимися при разложении азотной кислоты при нагревании.

Аналогичная изомеризация полиненасыщенных метиленразделенных высших жирных кислот протекает в более мягких условиях и приводит к сопряженным полиеновым кислотам. Это связано с большей подвижностью атома водорода метиленовой группы, расположенной между двойными связями. Подобного типа изомеризация полиеновых кислот при высокой температуре может протекать и в отсутствие щелочи.
Замещенные жирные кислоты. Такие кислоты встречаются редко; тем не менее известны соединения, содержащие в качестве заместителей гидрокси- и эпоксигруппы.
В жирах и маслах, не подвергавшихся окислению, гидроксикислоты содержатся редко и в небольших количествах. Исключением является касторовое масло, среди кислот которого до 94% приходится на рицинолевую ( 12-гидроксиолеиновую) кислоту, а также содержится 9,10-дигидроксистеариновая кислота. Благодаря такому составу касторовое масло обладает эмульгирующей способностью. Поэтому гидрогенизированное касторовое масло используется в приготовлении основ для мазей.
Высшие жирные кислоты, содержащие гидроксильную группу, входят в состав липидов головного мозга, липидов молока, некоторых растений и микроорганизмов. В липидах головного мозга содержатся а-гидроксикислоты С14—С26 как с четным, так и нечетным числом атомов углерода. Например, в состав цереброзидов (см. 37.3.3) входят гидрокси нервоновая (2-гидрокси- тетракозен-9-овая) и цереброновая (2-гидрокситетракозановая) кислоты.
Значительно чаще гидрокси кислоты встречаются в составе растительных восков. В них содержатся такие гидрокси кислоты насыщенного ряда, как сабиновая (12-гидроксилауриновая) и юниперовая (16-гидроксипальмити- новая) кислоты. В восках обнаружены также а-гидроксикислоты с разветвленной цепью атомов углерода.

Способ I. В сокращенном виде указывается число атомов углерода в углеводородной цепи и через двоеточие число двойных связей; положение двойных связей обозначается цифрами с указанием цис- или транс-конфигурации латинскими буквами с или t соответственно.
Поскольку природные высшие жирные кислоты имеют в основном цис-конфигурацию двойной связи, буквы с и t в сокращенных названиях

2.Заменимые, незаменимые высшие жирные кислоты, содержание в организме, витамин F.
Ненасыщенные жирные кислоты – олеиновая, линолевая, линоленовая и арахидоновая – называются незаменимыми, т. к. они не синтезируются в организме человека и должны поступать с пищей в количестве 5 г в сутки (с растительными маслами).
Особую роль в организме играет арахидоновая кислота как предшественник простагландинов. Ненасыщенные кислоты способствуют снижению содержания в крови холестерина и уменьшают опасность заболевания атеросклерозом.
Насыщенные жирные кислоты синтезируются в организме ферментативным путем из уксусной кислоты. Простые липиды, в состав которых входят преимущественно насыщенные кислоты – имеют твердую консистенцию и называются жирами, а триациглицерины, содержащие ненасыщенные кислоты – жидкие и называются маслами.
Некоторые биологически важные полиеновые кислоты. Полиненасыщенные высшие жирные кислоты (в медицинской литературе для них принята аббревиатура ПНВЖК) играют исключительно важную роль в жизнедеятельности организма.
Природные источники важнейших представителей полиеновых кислот приведены в табл. 3.
К полиненасыщенным высшим жирным кислотам относятся (см. табл. 2):

Таблица 3. Полиненасыщенные высшие жирные кислоты

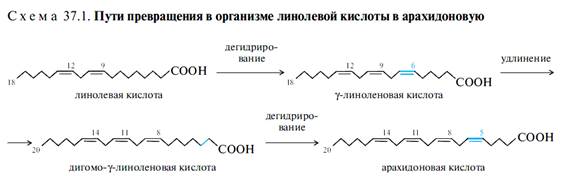
Полиеновые кислоты метаболически родственны. В организме они могут образовываться с помощью реакций дегидрирования и удлинения цепи. Так, арахидоновая кислота может быть синтезирована в организме из линолевой кислоты (схема 37.1), эйкозапентаеновая — из линоленовой кислоты. Сами же линолевая и линоленовая кислоты в организме не синтезируются, но они необходимы для нормального развития животных организмов. Поэтому кислоты такого типа были названы незаменимыми.
Считалось, что большинство функций незаменимых жирных кислот — линолевой, линоленовой и их более длинноцепочечных гомологов — связано с их окисленными метаболитами, получившими обобщенное название «оксилипины», а сами кислоты лишь обеспечивают целостность мембраны и оптимальный уровень ненасыщенности тканевых липидов. Позднее было показано, что высшие жирные кислоты непосредственно участвуют во многих клеточных процессах.
Перечисленные выше и родственные им полиненасыщенные кислоты условно называют «витамином F».
Линолевая и а-линоленовая кислоты являются компонентами растительных масел. γ-Линоленовая кислота значительно реже встречается в растительном мире. Однако она имеет не менее важное значение для жизнедеятельности и образуется в организме в качестве промежуточного продукта при превращении линолевой кислоты в арахидоновую (см. схему 37.1). Поступление ее с пищей служит дополнительным источником исходного продукта для получения арахидоновой кислоты.
Арахидоновая кислота является одной из наиболее значимых полиеновых кислот. С одной стороны, она обладает высокой биологической активностью и служит предшественником важных биологически активных веществ — эйкозаноидов, с другой стороны, является одной из важнейших жирных кислот, входящих в состав фосфолипидов. Арахидоновая кислота практически не содержится в липидах растений, а в организме животных образуется преимущественно из линолевой кислоты (см.схему 37.1).
Арахидоновую кислоту иногда относят (вместе с линолевой и линоленовой) к незаменимым кислотам, хотя она может быть синтезирована и в организме.

При поступлении в организм эйкозапентаеновой кислоты она включается в фосфолипиды мембран и после действия фосфолипазы служит основным субстратом для синтеза эйкозаноидов, оказывающих существенное влияние на регуляцию многих физиологических процессов, например на свертывание крови.
Докозагексаеновая кислота в большом количестве содержится в мозге, особенно в сером веществе (25—35% от общего количества всех жирных кислот фосфолипидов), в нервной ткани, в фоторецепторах сетчатки глаза. Дефицит этой кислоты приводит к нарушениям в функционировании мозга и нервной системы.
Пищевое поступление рыбьего жира, как источника ω-3 кислот, способствует нормализации баланса холестерина в крови, а именно понижению уровня липопротеинов низкой плотности и повышению уровня «хорошего» холестерина (липопротеинов высокой плотности), снижает свертываемость крови и улучшает ее реологические свойства.

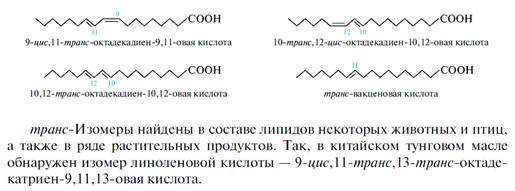
транс-Ненасыщенные высшие жирные кислоты. В последнее время появилось много информации об обнаружении и биологическом действии транс-форм ненасыщенных высших жирных кислот. Первые сведения по содержанию транс-изомеров были известны достаточно давно, транс-изомеры были обнаружены в молочных продуктах, где преобладающей является 9-цис,11-транс-октадекадиен-9,11-овая кислота и в небольших количествах содержатся 10-транс,12-цис- и 10-транс, 12-транс-октадекадиеновые кислоты. В процессе биогидрирования ненасыщенных жирных кислот, происходящего в анаэробной среде, например в рубце (передней части желудка) жвачных животных, образуется транс-вакценовая (транс-октадецен-11-овая) кислота.

3.Строение и свойства нейтральных липидов(жиры и масла), их консистенция, гидролиз.







4.Структура фосфолипидов, реакции гидролиза, образования, их биологическое значение.
К сложным относятся липиды, при пиролизе которых, кроме высших жирных кислот и высших спиртов, образуются и другие вещества, например фосфорная кислота, углеводы. Сложные липиды трудно классифицировать однозначно, так как они содержат группировки, позволяющие относить их одновременно к нескольким группам .
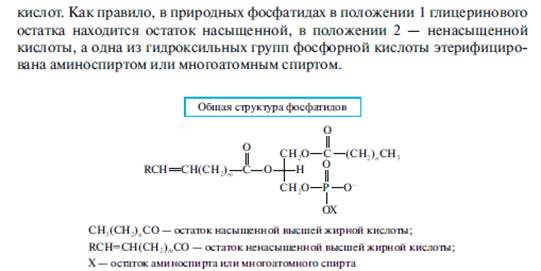
Отличительным признаком фосфолипидов является наличие в их структуре остатка фосфорной кислоты. По этому признаку к фосфолипидам причисляются глицерофосфолипиды, а также некоторые сфинголипиды .
Фосфолипидам принадлежит основная роль в построении и функционировании клеточных мембран. С участием фосфолипидных фрагментов мембран осуществляется ряд важнейших биохимических клеточных процессов. Мембранные липиды принимают участие в транспортировке ионов и различных молекул через стенки клетки.
Фосфолипиды входят в состав ряда лекарственных средств, применяемых для лечения и профилактики сердечно-сосудистых заболеваний, заболеваний нервной системы, печени.
Общим структурным фрагментом всех глицерофосфолипидов служит глицерофосфат, который содержит один асимметрический атом углерода С-2 и поэтому может существовать в виде двух стереоизомеров. Природные глицерофосфолипиды имеют одинаковую конфигурацию и являются производными L-глицеро-З-фосфата.
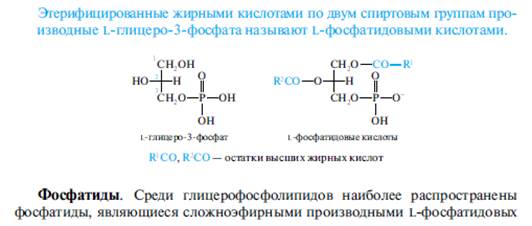
Помимо фосфолипидов с двумя ацильными остатками, во многих природных объектах в небольших количествах содержатся также производные глицерофосфата, имеющие всего лишь один остаток высшей жирной кислоты в положении 1. Они образуются в клетке под действием фосфолипаз и носят общее название лизофосфолипиды.
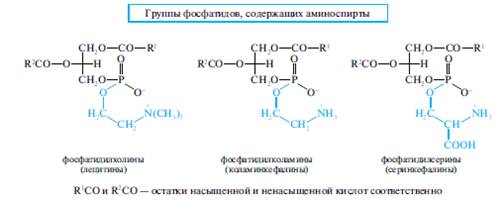
В зависимости от строения аминоспиртафосфатиды подразделяются на:
- фосфатидилхолины (аминоспирт— холин);
- фосфатидилэтаноламины, или фосфатидилколамины (аминоспирт — этаноламин, называемый также коламином);
- фосфатидилсерины (аминоспирт — серии).
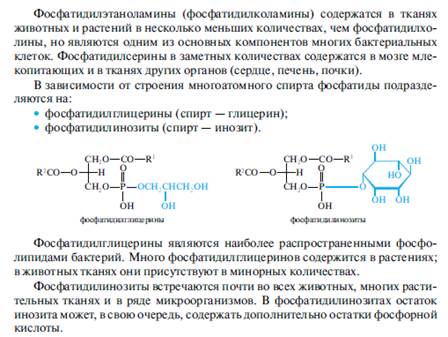

Плазмалогены являются альдегидогенными липидами, поскольку в результате их гидролиза в кислой среде образуются высшие жирные альдегиды (плазмали).
Плазмалогены обнаружены в тканях и органах всех животных независимо от уровня их организации. В достаточно высокой концентрации они присутствуют также в организме человека, где составляют 22% от общего количества фосфолипидов. Особенно велико их содержание в нервной ткани, головном мозге (белое вещество, мозговая оболочка), сердечной мышце, надпочечниках. В меньшей степени они представлены в микроорганизмах и растениях.


Образование (синтез) фосфолипидов. Благодаря этому пути холин и этаноламин используются повторно и не катаболизируют. Активация холина(или этаноламина)происходит через промежуточное образование фосфорилированных производных с последующим присоединением ЦМФ. В следующей реакции фосфохолин (или фосфозтаноламин) переносится на ДАГ. Этот путь особенно характерен для легких и кишечника, но идет и в других тканях.
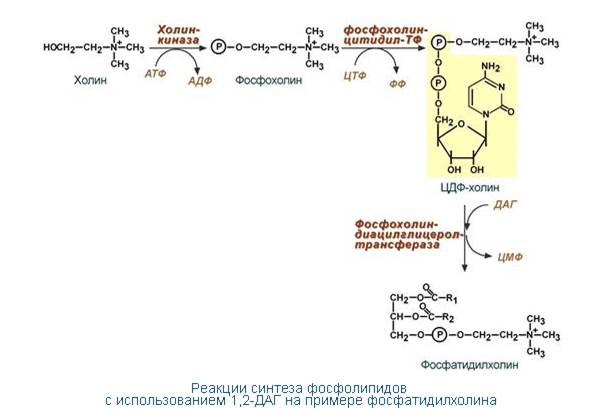
( ДАГ – диацилглицерин)
5.Соединения стероидной природы: холестерин, желчные кислоты.
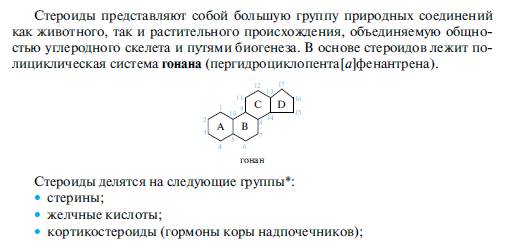

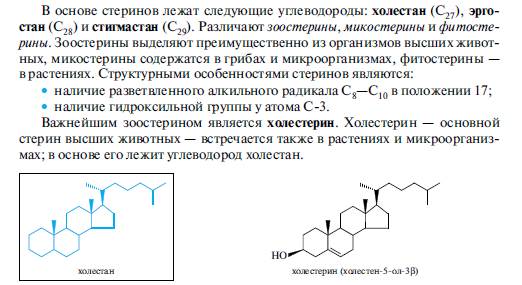
В животных организмах холестерин содержится в свободном виде или в виде сложных эфиров с карбоновыми кислотами, в том числе и с высшими. Холестерин или его сложные эфиры участвуют в построении клеточных мембран. Основное количество холестерина (-80%) человеческий организм синтезирует изацетилкофермента А и лишь около 20% получает с пищей. Эндогенный и алиментарный холестерин используется организмом для синтеза гормонов и желчных кислот. Излишек холестерина выводится из организма главным образом через пищеварительный тракт, в небольшом количестве холестерин экскретируется через сальные железы. Нарушение обмена холестерина приводит к отложению его на стенках кровеносных сосудов, что является одной из причин атеросклероза.
Получают холестерин из спинного мозга животных и из ланолина (жироподобного вещества, смываемого с овечьей шерсти), используют для синтеза стероидных гормонов и витамина D3.
| <== предыдущая лекция | | | следующая лекция ==> |
| Венчурное финансирование | | | Элементарный состав белков. |
Дата добавления: 2021-04-21; просмотров: 3355;
Поиск по сайту
Узнать еще
- A) Кремниевые кислоты и их соли
- I. Товар и его свойства.
- L-Аминокислоты ОРГАНИЗМА
- Акты применения норм права: понятие, структура, классификация, эффективность. Отличие правоприменительных актов от нормативно-правовых
- АЛГОРИТМ И ЕГО СВОЙСТВА. Определение алгоритма
- Амиды угольной кислоты
- АТС и его эксплуатационные свойства.
- БИОСИНТЕЗ БЕЛКА. ГЕНЕТИЧЕСКИЙ КОД И ЕГО СВОЙСТВА.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
