Функции нервной системы
Нервная система в организме человека выполняет следующие функции:
1. Обеспечивает взаимосвязь между органами и системами путем быстрой и точной передачи информации и ее интеграции.
2. Обеспечивает функционирование организма как единого целого и его взаимодействие с внешней средой.
3. Осуществляет прием и анализ разнообразных сигналов внешней и внутренней среды и формирует ответные реакции.
4. Осуществляет следующие психические функции:
— осознание сигналов окружающего мира,
— их запоминание,
— принятие решения и организация целенаправленного поведения,
— абстрактное мышление,
— речь.
2.2. Общий план строения и классификация нервной системы
Вся нервная система построена из нервной ткани, в состав которой входят высокоспециализированные нервные клетки, называемые нейронами и вспомогательные клетки — нейроглии.
Топографически нервную систему человека подразделяют на центральную и периферическую. К центральной нервной системе относят спинной и головной мозг. Периферическая нервная система образована нервными узлами (спинно-мозговыми, черепными и вегетативными), нервами (31 пара спинно-мозговых и 12 пар черепных) и нервными окончаниями, рецепторами (чувствительными) и эффекторами. Каждый нерв состоит из нервных волокон, миелинизированных и немиелинизированных.
По анатомо-функциональной классификации единую нервную систему также условно подразделяют на две части: соматическую (цереброспинальную) и вегетативную (автономную). Соматическая нервная система обеспечивает иннервацию главным образом тела (сому), кожи, скелетных мышц. Этот (соматический) отдел нервной системы устанавливает взаимоотношения с внешней средой, воспринимает ее воздействия (прикосновение, осязание, боль, температуру), формирует осознанные (управляемые сознанием) сокращения скелетных мышц (защитные и другие движения).
Вегетативная (автономная) нервная система иннервирует все внутренние органы (пищеварения, дыхания, мочеполовой аппарат), железы, гладкую мускулатуру органов, сердце, регулирует обменные процессы, рост и размножение.
Автономную (вегетативную) нервную систему на основании строения, топографии вегетативных ядер в спинном и головном мозге, а также особенностей функции, подразделяют на симпатическую и парасимпатическую части. Обе эти части вегетативной нервной системы действуют на одни и те же внутренние органы, не противоборствуя, а создавая более оптимальный режим их работы. В зависимости от жизненных обстоятельств, от величины функциональных нагрузок вегетативная нервная система или усиливает функции тех или иных внутренних органов, включая работу сердца, или ослабляет их. При этом в каждый момент в соответствии с потребностями организма большую активность в отношении внутренних органов проявляет или симпатическая, или парасимпатическая части вегетативной нервной системы. Что касается остальных органов и тканей (опорно-двигательного аппарата, кожи с их структурными элементами, стенок сосудов и некоторых других), то все обменные процессы в них регулирует симпатическая часть вегетативной нервной системы.
Координацию работы всех отделов вегетативной нервной системы осуществляют гипоталамус промежуточного мозга и кора большого мозга.
2.3. Нейрон как структурная и функциональная единица
нервной системы
Нейрон — это основная структурная и функциональная единица нервной системы. Нейроном называют нервную клетку с отростками.
В нем различают тело клетки, или сому, один длинный, мало ветвящийся отросток — аксон — и много (от 1 до 1000) коротких, сильно ветвящихся отростков — дендритов (рис. 3А). Длина аксона достигает метра и более, его диаметр колеблется от сотых долей микрона до 10 мкм; длина дендрита может достигать 300 мкм, а его диаметр — 5 мкм.
Аксон, выходя из сомы клетки, постепенно суживается, от него отходят отдельные отростки — коллатерали. На протяжении первых
50—100 мкм от тела клетки аксон не покрыт миелиновой оболочкой. Прилегающий к нему участок тела клетки называют аксонным холмиком. Участок аксона, не покрытый миелиновой оболочкой, вместе с аксонным холмиком называют начальным сегментом аксона. Эти участки отличаются рядом морфологических и функциональных особенностей.
По дендритам возбуждение поступает от рецепторов или других нейронов к телу клетки, а аксон передает возбуждение от тела нейрона к другому или рабочему органу. На дендритах имеются боковые отростки (шипики), которые увеличивают их поверхность и являются местами наибольших контактов с другими нейронами. Конец аксона сильно ветвится, один аксон может контактировать с 5 тыс. нервных клеток и создавать до 10 тыс. контактов. Место контакта одного нейрона с другим получило название синапса (от греческого слова «синапто» — контактировать). По внешнему виду синапсы имеют форму пуговки, луковицы, петли и др. Количество синаптических контактов неодинаково на теле и отростках нейрона и очень вариабельно в различных отделах центральной нервной системы. Тело нейрона на 38 % покрыто синапсами, и их насчитывают до 1200—1800 на одном нейроне. Много синапсов на дендритах и шипиках, их количество невелико на аксонном холмике. Все нейроны центральной нервной системы соединяются друг с другом в основном в одном направлении: разветвления аксона одного нейрона контактируют с телом клетки и дендритами другого нейрона.
Тело нервной клетки в различных отделах нервной системы имеет разную величину (диаметр его колеблется от 4 до 130 мкм) и форму (округлую, уплощенную, многоугольную, овальную). Оно покрыто сложно устроенной мембраной и содержит органеллы, свойственные любой другой клетке. В теле находятся ядро с одним или несколькими ядрышками, митохондрии, рибосомы, аппарат Гольджи, эндоплазматическая сеть и др. Характерной особенностью строения нервной клетки является наличие гранулярного ретикулума с большим количеством рибосом и нейрофибрилл. С рибосомами в нервных клетках связывают высокий уровень обмена веществ, синтез белка и РНК. Нейрофибриллы представляют собой тончайшие волоконца, пересекающие тело клетки во всех направлениях и продолжающиеся в отростки и участвующие в проведении нервных импульсов (рис. 3Б).
В ядре содержится генетический материал — дезоксирибонуклеиновая кислота (ДНК), которая регулирует состав РНК сомы нейрона. РНК в свою очередь определяет количество и тип белка, синтезируемого в нейроне.
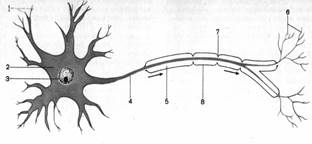
А

Б
Рис. 3. Структура нервной клетки:
А — Строение нервной клетки: 1 — дендрит, 2 — тело клетки,
3 — ядро,4 — аксон, 5 — миелиновое волокно, 6 — ветви аксона,
7 — перехват, 8 — неврилемма;
Б — нейрофибриллы в двигательной клетке спинного мозга
Нейроны различают по строению и функции. По строению (в зависимости от количества отходящих от тела клетки отростков) различают униполярные (с одним отростком), биполярные (с двумя отростками) и мультиполярные (с множеством отростков) нейроны.
По функциональным свойствам выделяют афферентные (или центростремительные) нейроны, несущие возбуждение от рецепторов в центральную нервную систему, эфферентные, двигательные, мотонейроны (или центробежные), передающие возбуждение из центральной нервной системы к иннервируемому органу, и вставочные, контактные или промежуточные нейроны, соединяющие между собой афферентные и эфферентные пути.
Афферентные нейроны относятся к униполярным, их тела лежат в спинно-мозговых ганглиях. Отходящий от тела клетки отросток Т-образно делится на две ветви, одна из которых идет в центральную нервную систему и выполняет функцию аксона, а другая подходит к рецепторам и представляет собой длинный дендрит.
Большинство эфферентных и вставочных нейронов относится к мультиполярным. Мультиполярные вставочные нейроны в большом количестве располагаются в задних рогах спинного мозга, находятся и во всех других отделах центральной нервной системы. Они могут быть и биполярными, как например, нейроны сетчатки, имеющие короткий ветвящийся дендрит и длинный аксон. Мотонейроны располагаются в основном в передних рогах спинного мозга.
Нейроглия находятся между нейронами и составляет межклеточное вещество нервной ткани. В состав глии входят клетки с отростками и без отростков и волокна, являющиеся либо отростками клеток, либо самостоятельными образованиями, называемыми глиофибриллами. Клетки нейроглии в нервной системе располагаются по-разному: находятся в белом веществе мозга, на значительном расстоянии покрывают клетки спинного мозга, идут вдоль определенной стороны нервной клетки, являясь спутниками нервных клеток. В некоторых отделах мозга они располагаются вдоль сосудов и обладают способностью к фагоцитозу. Установлено, что нейроглия имеет отношение к обмену веществ в нервной ткани. Нейроглия располагаясь вдоль сосудов, обеспечивает питание нейронов. Клетки нейроглии выстилают спинно-мозговой канал и желудочки мозга и участвуют в секреции спинно-мозговой жидкости: клетки, обладающие длинными отростками, участвуют в образование опорных структур вокруг тел нейронов; клетки, лишенные отростков, соединяясь друг с другом, образуют оболочки нервных волокон; мелкие клетки образуют небольшие скопления в головном мозге и выполняют защитные функции. Некоторые клетки нейроглии выделяют вещества, влияющие на состояние возбудимости нервных клеток. Отмечено, что при разных психических состояниях изменяется секреция этих клеток. С функциональным состоянием нейроглии связывают длительные следовые процессы в центральной нервной системе.
На ранних стадиях развития нервная клетка характеризуется большой величиной ядра, которое окружено небольшим количеством цитоплазмы. Такая клетка носит название аполярного нейробласта. В процессе развития с увеличением размеров клетки уменьшается относительный объем ядра. На третьем месяце внутриутробного развития в аксоплазме появляются нейрофибриллы, и одновременно начинается рост аксона нервной клетки. Аксон растет по направлению к периферии вплоть до конечного органа мышцы или железы. Во время развития аксона происходит его погружение в шванновскую клетку и образование миелиновой оболочки.
Растущий аксон на конце имеет колбу роста. Синапс центральной нервной системы образуется в результате контакта колбы роста одного нейрона с телом другого. Колба роста превращается в пресинаптическое образование. На первом этапе развития синапса дифференцируются мембраны, затем в пресинаптическом отделе образуются митохондрии и везикулы, количество которых быстро увеличивается. Постепенно увеличивается синаптическая щель и утолщаются мембраны синапса. Функциональная деятельность нейрона начинается с того момента, когда аксон достигает органа. Проведение возбуждения в нейронах центральной нервной системы обнаружено с момента образования синапса со всеми его компонентами. Дендриты вырастают значительно позже аксона. Сначала на противоположном аксону полюсе клетки появляется верхушечный дендрит в виде простого выроста аксоплазмы, вследствие чего нейробласт становится биполярным. Затем вырастают дендриты со всех сторон, и нейробласт становится мультиполярным. Способность проводить возбуждение у дендрита появляется значительно позже, чем у аксона (аксон функционирует во внутриутробном периоде развития ребенка, а дендрит — после рождения), В процессе развития увеличивается число ветвлений дендрита. Шипики на дендритах появляются после рождения ребенка. В коре больших полушарий их количество возрастает вместе с увеличением числа условно-рефлекторных связей.
2.4. Строение, свойства и возрастные изменения
нервных волокон
Нервным волокном называют отросток нервной клетки, покрытый оболочками. Центральную часть любого отростка нервной клетки (аксона или дендрита) называют осевым цилиндром. Осевой цилиндр располагается в аксоплазме и состоит из тончайших волокон — нейрофибрилл и покрыт оболочкой — аксолеммой. При рассмотрении под электронным микроскопом установлено, что каждая нейрофибрилла состоит из еще более тонких волокон разного диаметра, имеющих трубчатое строение. Трубочки диаметром до 0,03 мкм называют нейротубулями, а диаметром до 0,01 мкм — нейрофиламентами. По нейротубулям и нейрофиламентам поступают к нервным окончаниям вещества, образующиеся в теле клетки и служащие для передачи нервного импульса.
В аксоплазме содержатся митохондрии, количество которых особенно велико в окончаниях волокон, что связывают с передачей возбуждения с аксона на другие клеточные структуры. В аксоплазме мало рибосом и РНК, чем объясняется низкий уровень обмена веществ в нервном волокне.
Аксон покрыт миелиновой оболочкой до места его разветвления у иннервируемого органа, которая располагается вдоль осевого цилиндра не сплошной линией, а сегментами длиной 0,5—2 мм. Пространство между сегментами (1—2 мкм) называют перехватом Ранвье. Миелиновая оболочка образуется шванновскими клетками путем их многократного обкручивания вокруг осевого цилиндра. Каждый ее сегмент образован одной шванновской клеткой, скрученной в сплошную спираль.
В области перехватов Ранвье миелиновая оболочка отсутствует, и концы шванновских клеток плотно прилегают к аксолемме. Наружная мембрана шванновских клеток, покрывающая миелин, образует самую верхнюю оболочку нервного волокна, которую называют шванновской оболочкой или неврилеммой. Шванновским клеткам придают особое значение, их считают клетками-спутниками, которые дополнительно обеспечивают обмен веществ в нервном волокне. Они принимают участие в процессе регенерации нервных волокон.
Различают мякотные, или миелиновые, и безмякотные, или безмиелиновые, нервные волокна. К миелиновым относят волокна соматической нервной системы и некоторые волокна вегетативной нервной системы. Безмякотные волокна отличаются тем, что в них не развивается миелиновая оболочка и их осевые цилиндры покрыты только шванновскими клетками (шванновской оболочкой). К ним относится большинство волокон вегетативной нервной системы.
Свойства нервных волокон. В организме возбуждение проводится по нервам, в состав которых входит большое количество различных по строению и функции нервных волокон.
Основные свойства нервных волокон заключаются в следующем: связь с телом клетки, высокая возбудимость и лабильность, невысокий уровень обмена веществ, относительная неутомляемость, большая скорость проведения возбуждения (до 120 м/с). Миелинизация нервных волокон осуществляется в центробежном направлении, отступая несколько микрон от тела клетки к периферии нервного волокна. Отсутствие миелиновой оболочки ограничивает функциональные возможности нервного волокна. Реакции возможны, но они диффузные и слабо координированы. По мере развития миелиновой оболочки возбудимость нервного волокна постепенно повышается. Раньше других начинают миелинизироваться периферические нервы, затем волокна спинного мозга, стволовой части головного мозга, мозжечка и позже — больших полушарий головного мозга. Миелинизация спинно-мозговых и черепно-мозговых нервов начинается на четвертом месяце внутриутробного развития. Двигательные волокна покрыты миелином к моменту рождения. Большинство смешанных и центростремительных нервов миелинизируются к трем месяцам после рождения, некоторые — к трем годам. Проводящие пути спинного мозга хорошо развиты к моменту рождения и почти все миелинизированы. Не заканчивается миелинизация только пирамидных путей. Скорость миелинизации черепно-мозговых нервов различна; большинство из них миелинизируются к 1,5—2 годам. Миелинизация нервных волокон головного мозга начинается во внутриутробном периоде развития и заканчивается после рождения. Несмотря на то, что к трем годам в основном заканчивается миелинизация нервных волокон, рост в длину миелиновой оболочки и осевого цилиндра продолжается и после трехлетнего возраста.
2.5. Строение синапса. Механизм передачи возбуждения
в синапсах
Синапс состоит из пресинаптического и постсинаптического отделов, между которыми имеется небольшое пространство, получившее название синоптической щели (рис. 4).
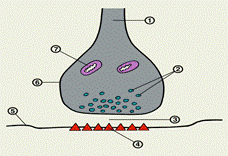 |
Рис. 4. Межнейрональный синапс:
1 — аксон; 2 — синаптические пузырьки; 3 — синаптическая щель;
4 — хеморецепторы постсинаптической мембраны; 5 — поссинаптическая мембрана; 6 — синаптическая бляшка; 7 — митохондрия
Благодаря электронно-микроскопической технике исследования обнаружены синаптические контакты между различными образованиями нейронов. Синапсы, образованные аксоном и телом (сомой) клетки, называют аксосоматическими, аксоном и дендритом аксодендритическими. В последнее время изучены контакты между аксонами двух нейронов — они получили название аксо-аксональных синапсов. Соответственно контакты между дендритами двух нейронов называют дендро-дендритическими синапсами.
Синапсы между окончанием аксона и иннервируемым органом (мышцей) получили название нервно-мышечных синапсов или концевых пластинок. Пресинаптический отдел синапса представлен конечной веточкой аксона, которая на расстоянии 200—300 мкм от контакта теряет миелиновую оболочку. В пресинаптическом отделе синапса содержится большое количество митохондрий и пузырьков (везикул) округлой или овальной формы размером от 0,02 до 0,05 мкм. В везикулах содержится вещество, способствующее передаче возбуждения с одного нейрона на другой, которое называют медиатором. Везикулы концентрируются вдоль поверхности пресинаптического волокна, находящейся против синаптической щели, ширина которой равна 0,0012—0,03 мкм. Постсинаптический отдел синапса образуется мембраной сомы клетки или ее отростков, а в концевой пластинке — мембраной мышечного волокна. Пресинаптическая и постсинаптическая мембраны имеют специфические особенности строения, связанные с передачей возбуждения: они несколько утолщены (их диаметр около 0,005 мкм). Длина этих участков составляет 150—450 мкм. Утолщения могут быть сплошными и прерывистыми. Постсинаптическая мембрана у некоторых синапсов складчатая, что увеличивает поверхность соприкосновения ее с медиатором. Аксо-аксональные синапсы имеют строение, подобное аксо-дендритическим, в них везикулы располагаются в основном с одной (пресинаптической) стороны.
Механизм передачи возбуждения в концевой пластинке. В настоящее время представлено много доказательств химической природы передачи импульса и изучен ряд медиаторов, т. е. веществ, способствующих передаче возбуждения с нерва на рабочий орган или с одной нервной клетки на другую.
В нервно-мышечных синапсах, в синапсах парасимпатической нервной системы, в ганглиях симпатической нервной системы, в ряде синапсов центральной нервной системы медиатором является ацетилхолин. Эти синапсы названы холинэргическими.
Обнаружены синапсы, в которых передатчиком возбуждения является адреналиноподобное вещество; они названы адреналеэгическими. Выделены и другие медиаторы: гаммааминомасляная кислота (ГАМК), глютаминовая и др.
Прежде всего было изучено проведение возбуждения в концевой пластинке, так как она более доступна для исследования. Последующими экспериментами было установлено, что в синапсах центральной нервной системы осуществляются аналогичные процессы. Во время возникновения возбуждения в пресинаптической части синапса увеличивается количество везикул и скорость их движения. Соответственно увеличивается количество ацетилхолина и фермента холинацетилазы, способствующего его образованию. При раздражении нерва в пресинаптической части синапса одновременно разрушается от 250 до 500 везикул, соответственно выделяется в синаптическую щель такое же количество квантов ацетилхолина. Это связано с влиянием, ионов кальция. Его количество в наружной среде (со стороны щели) в 1000 раз больше, чем внутри пресинаптического отдела синапса. Во время деполяризации увеличивается проницаемость пресинаптической мембраны для ионов кальция. Они входят в пресинаптическое окончание и способствуют вскрытию везикул, обеспечивая выход ацетилхолина в синаптическую щель.
Выделившийся ацетилхолин диффундирует к постсинаптической мембране и действует на участки, особенно к нему чувствительные,— холинорецепторы, вызывая возбуждение в постсинаптической мембране. На проведение возбуждения через синаптическую щель затрачивается около 0,5 м/с. Это время получило название синаптической задержки. Оно слагается из времени, в течение которого происходит освобождение ацетилхолина, диффузии его от пресинаптической мембраны
к постсинаптической и воздействия на холинорецепторы. В результате действия ацетилхолина на холинорецепторы открываются поры постсинаптической мембраны (мембрана разрыхляется и становится на короткое время проницаемой для всех ионов). При этом в постсинаптической мембране возникает деполяризация. Одного кванта медиатора достаточно для того, чтобы слабо деполяризовать мембрану и вызвать потенциал амплитудой 0,5 мВ. Такой потенциал называют миниатюрным потенциалом концевой пластинки (МПКП). При одновременном освобождении 250—500 квантов ацетилхолина, т. е. 2,5—5 млн молекул, наступает максимальное увеличение числа миниатюрных потенциалов.
Дата добавления: 2016-09-06; просмотров: 7177;
Поиск по сайту
Узнать еще
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I.2. Антигены системы АВ0. Генетика. Структура
- I.2.1 ПЕРВЫЙ ЗАКОН НЬЮТОНА. ИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА.
- I.2.1 ПОЛНАЯ И ВНУТРЕННЯЯ ЭНЕРГИЯ СИСТЕМЫ. ТЕПЛОТА И РАБОТА
- I.3. Антитела системы АВ0
- I.5.4 НЕИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА. СИЛЫ ИНЕРЦИИ
- II. Митохондрии (строение и функции)

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
