Морфолого-анатомическая характеристика клубеньков в их онтогенезе
По способу образования клубеньки бобовых растений подразделяются на два типа: 1-й тип — клубеньки возникают при делении клеток перицикла (кор-неродного слоя), обычно расположенных против протоксилемы (первых по времени образования сосудов) — эндогенный тип образования клубеньков; 2-й тип — клубеньки происходят из коры корня в результате внедрения возбудителя в паренхимные клетки коры и эндодермы (внутреннего слоя первичной коры) — экзогенный тип образования клубеньков.
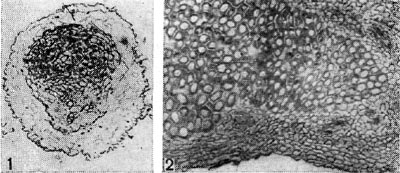
Рис. 153. Срезы через клубеньковую ткань бобовых растений: 1 — поперечный срез через клубенек гороха. Начало дифференциации сосудистой ткани в клубеньке (по Г. Бонду); г — продольный срез через клубенек чины. Увел. X 90.
В природе преобладает последний тип. Ткани центрального цилиндра корня принимают участие только в образовании сосудистой системы клубеньков как эндогенного, так и экзогенного типа.
Несмотря на различные взгляды на природу возникновения клубеньков экзо- и эндотипов, процесс развития их в основном одинаков. Однако ни тот, ни другой тип образования клубеньков ни в коем случае не следует отождествлять с процессом образования боковых корней, несмотря на то что существуют и отдельные черты сходства в их заложении. Так, формирование клубеньков и боковых корней происходит одновременно и к тому же в одной и той же зоне корня.
В то же время ряд особенностей развития боковых корней и клубеньков подчеркивает глубокие различия в типе их формирования. Боковые корни возникают в перицикле. С первых же моментов развития они связаны с центральным цилиндром главного корня, от которого ответвляются центральные цилиндры боковых корней,и возникают они всегда против луча первичной древесины. Формирование клубенька, в отличие от бокового корня, возможно в любом месте. В самом начале формирования клубеньковой ткани сосудистой связи с центральным цилиндром корня нет, она возникает позднее. Сосуды обычно формируются по периферии клубенька. Они связаны с сосудами корня через зону трахеид и имеют собственную эндодерму (рис. 152).
Различие в характере возникновения клубеньков и боковых корней особенно четко наблюдается у сераделлы, поскольку коровая ткань главного корня этого растения — место возникновения первых клубеньков — состоит из относительно небольшого слоя клеток и клубеньки становятся видимыми очень быстро после инфицирования корня бактериями. Они образуют сначала выступы уплощенной формы на корне, что позволяет отличить их от конических выступов боковых корней. Клубеньки отличаются от боковых корней и рядом анатомических признаков: отсутствием центрального цилиндра, корневых чехликов и эпидермиса, наличием значительного слоя коры, покрывающей клубенек.
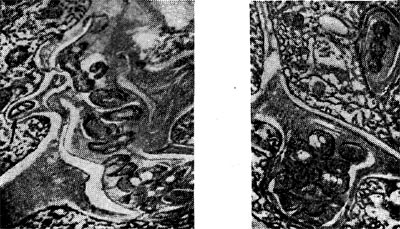
Рис. 154. Интерцеллюлярные инфекционные нити в виде кармана (справа) и факела (слева) с клубеньковыми бактериями. Стрелкой показана клетка бактерий, приготовившаяся к выходу в цитоплазму растительной клетки. Увел. X 20 000.
Формирование клубеньков (рис. 153, 1, 2) бобовых растений происходит в период, когда корень имеет еще первичную структуру. Оно начинается с деления коровых клеток, расположенных на расстоянии 2—3 слоев от концов инфекционных нитей. Слои коры, пронизанные инфекционными нитями, остаются без изменения. В то же время у сераделлы деление коровых клеток возникает непосредственно под инфицированным корневым волоском, а у гороха деление клеток отмечается только в предпоследнем слое коры.
Деление с образованием радиальной структуры ткани продолжается до внутренних коровых клеток. Происходит оно без определенного направления, беспорядочно, и в результате этого возникает меристема (система образовательных тканей) клубенька, состоящая из мелких зернистых клеток.
Разделившиеся клетки коры изменяются: ядра округляются и увеличиваются в размерах, особенно увеличиваются ядрышки. После митоза ядра расходятся и, не принимая первоначальной формы, вновь начинают делиться.
Возникает вторичная меристема. Вскоре в эндодерме и перицикле появляются признаки начинающегося деления, которое в прежних внешних клетках происходит главным образом тангентальными перегородками. Это деление распространяется, наконец, на общий меристе-матический комплекс, мелкие клетки которого вытягиваются, вакуоли исчезают, ядро заполняет большую часть клетки. Образуется так называемый первичный клубенек, в плазме клеток которого клубеньковые бактерии отсутствуют, поскольку они на данной стадии еще находятся внутри инфекционных нитей. В то время как образуется первичный клубенек, инфекционные нити многократно разветвляются и могут проходить или между клетками — интерцеллюлярно (рис. 154), или сквозь клетки — интрацеллюлярно — и вносить бактерии (рис. 155).
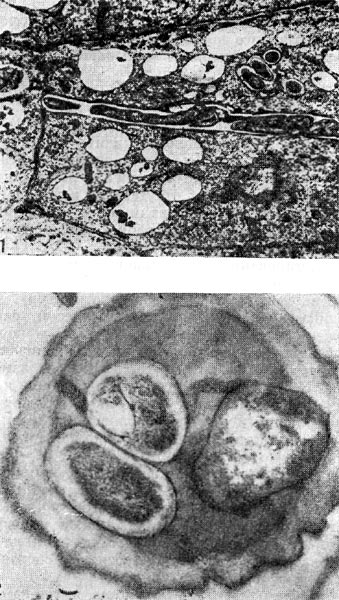
Рис. 155. Интрацеллюлярные инфекционные нити с клубеньковыми бактериями: 1 — продольный разрез (по Д. Гудчильду и Ф. Бергерсену), 2 — поперечный срез. Увел, х 35 000.
Межклеточные инфекционные нити вследствие активного размножения в них клубеньковых бактерий нередко приобретают причудливую форму — формируются в виде карманов (дивертикулов) или факелов (см. рис. 154).
Процесс передвижения инфекционных нитей из клетки в клетку не совсем ясен. По-видимому, инфекционные нити, как полагает канадский микробиолог Д. Джордан (19(53), блуждают в виде голых слизистых тяжей в межклеточных промежутках растительной ткани до тех пор, пока вследствие каких-то еще необъяснимых причин не начинают инвагинировать в цитоплазму примыкающих клеток.
В некоторых случаях инвагинация инфекционной нити происходит в одну, в некоторых случаях — в каждую соседнюю клетку. По этим инвагинированным трубчатым полостям (дивертикулам) перетекает заключенное в слизь содержимое нити. Наиболее активный рост инфекционных нитей происходит обычно вблизи ядра растительной клетки. Проникновение нити сопровождается перемещением ядра, которое продвигается к месту инфекции, увеличивается, меняет форму и дегенерирует. Подобная картина наблюдается при грибной инфекции, когда ядро нередко устремляется навстречу внедрившимся гифам, притягивается к повреждению как к месту наибольшей физиологической активности, вплотную придвигается к нити, разбухает и разрушается. По-видимому, это характерно для ответной реакции растения на инфекцию.
У однолетних растений инфекционные нити возникают обычно в первый период инфицирования корня, у многолетних — в течение длительного периода развития.
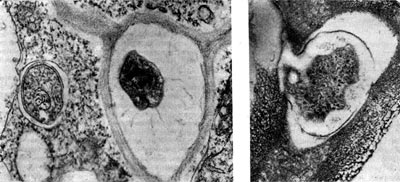
Рис. 156. Клетки клубеньковых бактерий в межклеточном пространстве (слева, по Д. Джордану) и в момент входа из межклеточного пространства в клетку растения (справа). Увел. X 60 000.
Бактерии могут высвобождаться из инфекционной нити в разное время и разными способами. Выход бактерий, как правило, весьма длительный процесс, особенно у многолетних растений. Обычно выход бактерий из инфекционной нити в цитоплазму растения-хозяина связывают с внутренним давлением, возникающим вследствие интенсивного размножения бактерий в нити и экскреции ими слизи. Иногда бактерии выскальзывают из нити группами, окруженными слизью инфекционной нити. в виде везикул (пузыревидных образований) (рис. 157). Поскольку везикулы не имеют оболочек, выход из них бактерий очень прост. В клетки растений клубеньковые бактерии могут попадать и поодиночке из межклеточных пространств (рис. 156).
Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина. Размножение их в этот период происходит делением перетяжкой (рис. 158). Основная масса бактерий размножается в цитоплазме клетки, а не в инфекционной нити. Зараженные клетки дают начало будущей бактероидной ткани.
Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. В момент митотического деления зараженных клеток клубеньковые бактерии могут скапливаться на двух противоположных полюсах материнской клетки и пассивно попадать в дочерние клетки. Каждая из незараженных клеток находится при этом под сильным стимулирующим воздействием клубеньковых бактерий и вследствие этого также делится. Благодаря такому энергично протекающему митотическому делению меристематических клеток осуществляется распространение клубеньковых бактерий в ткани клубенька и увеличение объема бактероидной области.
Инфицированная ткань, состоящая из плотно лежащих и активно делящихся клеток, имеет сначала форму усеченного конуса. В дальнейшем вследствие постепенного роста этого конуса и одновременного деления и развития меристематических клеток ткань клубенька разрастается, утрачивая конусовидность.
Таким образом, клубенек увеличивается сначала в результате радиального и тангенталь-ного деления коровых клеток, а затем за счет увеличения их размера и одновременного деления. После того как растительные клетки полностью заполнятся бактериями, митоз прекращается. Однако клетки продолжают увеличиваться в размере и часто сильно вытягиваются. Размер их в несколько раз больше, чем у неинфицированных растительных клеток, которые расположены между ними в бактероидной зоне клубенька.
Связь молодого клубенька с корнем бобового растения осуществляется благодаря сосудисто-волокнистым пучкам. Впервые сосудисто-волокнистые пучки наблюдал М. С. Воронин (1866). Время возникновения сосудистой системы в клубеньках различных видов бобовых растений различно. Так, у клубеньков сои начало развития сосудистых пучков совпадает с моментом проникновения клубеньковых бактерий в два слоя коровой паренхимы. С ростом клубенька проводящая система разрастается, разветвляется и окружает бактероидную область.
Параллельно с процессом дифференциации сосудистой системы идет формирование клубеньковой эндодермы из внешнего слоя первичного клубенька. Затем клубенек округляется, его периферийный клеточный слой окружается клубеньковой корой.
Корневой эпидермис разрывается, а клубенек продолжает развиваться и увеличиваться в размерах.
С помощью светового микроскопа на продольных срезах зрелых клубеньков обычно четко выделяются 4 характерные зоны тканевой дифференциации: кора, меристема, бакте-роидная зона и сосудистая система. Все ткани клубенька дифференцируются в акропетальной последовательности, так как новые клетки закладываются меристемой.
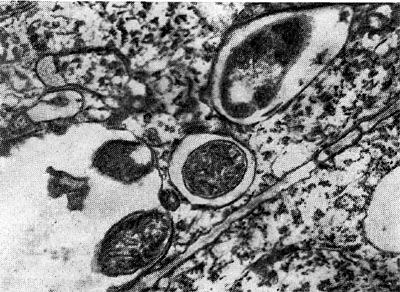
Рис. 157. Выход клубеньковых бактерий в ткань корня из везикулоподобных образований инфекционных нитей (по Гудчильду и Бергерсену).
Клубеньковая кора — оболочка клубенька, выполняющая защитную функцию. Кора состоит из нескольких рядов незараженных паренхимных клеток, величина и особенности которых различны у разных бобовых культур. Чаще всего клетки коры имеют вытянутую форму и крупнее по сравнению с другими клетками клубенька.
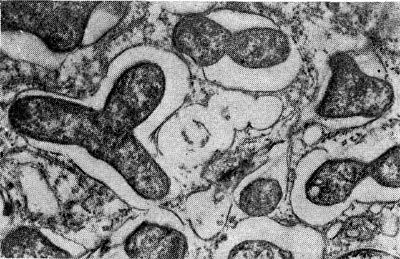
Рис. 158. Размножение клубеньковых бактерий в клубеньке чины. Увел. X 28 000.
В коре клубеньков многолетних деревянистых видов часто встречаются клетки с опробковевшими оболочками, содержащие смолы, танин, дубильные вещества.
Клубеньковая меристема расположена под клетками коры и представляет собой зону интенсивно делящихся также незараженных клеток. Для меристемы клубенька характерны плотно расположенные, без межклетников, мелкие тонкостенные клетки неправильной формы. Клетки меристемы клубенька подобны клеткам других типов меристематической ткани (верхушки корня, верхушки стебля). Клетки клубеньковой меристемы содержат плотную, тонко гранулированную цитоплазму с рибосомами, телами Гольджи, протопластидами, митохондриями и другими структурами. Встречаются небольшие вакуоли. В центре цитоплазмы расположено крупное ядро с ядерной мембраной, порами и четко выраженным ядрышком. Функции меристематических клеток заключаются в формировании клеток клубеньковой коры, бактероидной области и сосудистой системы. В зависимости от расположения меристемы клубеньки имеют разнообразную форму: шаровидную (горох, фасоль, сераделла, арахис) или цилиндрическую (люцерна, вика, чина, акация, клевер) (рис. 159). Меристема, расположенная отдельными участками по периферии клубенька, приводит к образованию муфтооб-разных клубеньков у люпина.
Клубеньковая меристема функционирует долго, даже во время некроза клубеньков, когда они уже наполнены лизирующейся бактероидной массой и разрушенными растительными клетками.
Бактероидная зона клубенька занимает его центральную часть и составляет от 16 до 50% от общей сухой массы клубеньков. В первый период формирования клубенька она, по существу, является бактериальной зоной (рис. 160), так как заполнена клетками бактерий, находящихся в бактериальной, а не бактероидной стадии развития. Тем не менее принято, когда идет речь о зоне клубеньковой ткани, содержащей бактерии, называть ее бактероидной.
Бактероидная область клубенька состоит в основном из инфицированных клубеньковыми бактериями клеток и частично из смежных с ними неинфицированных клеток, заполненных пигментами, дубильными веществами, а к осени — крахмалом.
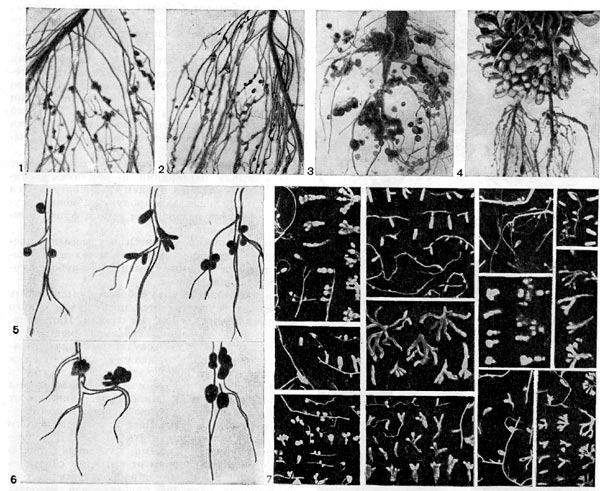
Ряс. 159. Форма клубеньков у бобовых растений: 1, 2 — гороха ; 3, 6 (справа) — люпина; 4 — арахиса (по Е. Швингхамеру); 5 — чины, клевера, вики, 6 (слева) — люцерны (по Я. Дэйту); 7 — разных видов дикорастущих бобовых растений (по А.Р. Вернеру, Н.Я. Гордиенко).
В клубеньках, образованных эффективными штаммами клубеньковых бактерий, средний относительный объем бактероидной зоны выше, чем в клубеньках, сформировавшихся при внедрении неэффективных штаммов.
В некоторых случаях объем бактероидной области достигает максимума в ранний период жизни клубенька и впоследствии остается относительно постоянным. Бактероидная зона пронизана густой сетью инфекционных нитей, а по периферии окружена сосудисто-волокнистыми пучками.
Форма бактероидов в клубеньках разных видов бобовых культур может быть разнообразной (табл. 44). Так, у вики, чины и гороха они двухветвистые или вильчатые. Для клевера и эспарцета преобладающая форма бактероидов шаровидная, грушеобразная, вздутая, яйцевидная, для нута округлая. Форма бактероидов фасоли, сераделлы, лядвенца и люпина практически палочковидная.
Бактероиды заполняют большую часть растительной клетки, за исключением центральной зоны ядра и вакуолей. Так, процент бактероидов в бактероидной зоне окрашенного в розовый цвет клубенька составляет 94,2 к общему числу клубеньковых бактерий. Клетки бактероидов в 3—5 раз больше клеток бактерий (рис. 161,1, 2).
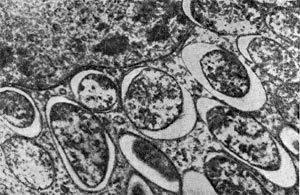
Рис. 160. Клетки клубеньковых бактерий вокруг ядра растительной клетки клубеньков вики. Увел. X 20 000.
Бактероиды клубеньковых бактерий представляют особый интерес в связи с тем, что они являются чуть ли не единственными обитателями клубеньков бобовых растений в период интенсивного связывания ими атмосферного азота. Отдельные исследователи считают бактероиды патологическими дегенеративными формами и не связывают процесс азотфиксации с бактероидной формой клубеньковых бактерий. Большинство исследователей находят, что бактероиды являются самыми жизнеспособными и активными формами клубеньковых бактерий и что фиксация азота атмосферы бобовыми растениями осуществляется только при их участии (рис. 162).
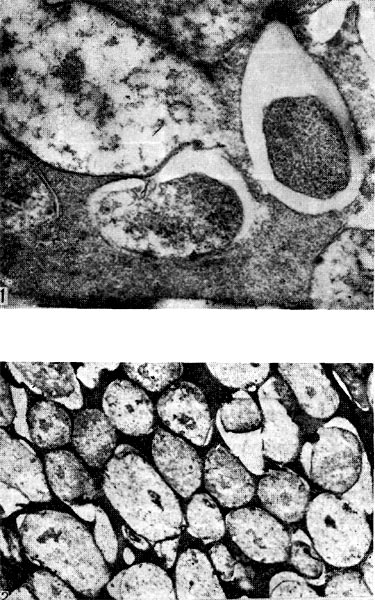
Рис. 161. Переход бактерий в бактероиды (1 — увел. Х25 000) и сформировавшаяся бактероидная ткань (2 — увел. X15 000) в клубеньках люцерны.
Сосудистая система клубенька обеспечивает связь между бактериями и растением-хозяином. По сосудистым пучкам транспортируются питательные вещества и продукты обмена. Сосудистая система развивается рано и функционирует длительное время.
Вполне сформировавшиеся сосуды имеют определенное строение: состоят из трахеид ксилемы, волокон флоэмы, ситовидных трубок и сопровождающих клеток.
В зависимости от вида бобовых культур связь клубецька осуществляется посредством одного или нескольких сосудистых пучков. Например, у гороха в основании клубенька имеется два дифференцированных сосудистых узла. Каждый из них обычно дважды дихотомически разветвляется, и в результате сквозь клубенек от места второго дихотомического разветвления проходит 8 пучков. Многие растения имеют лишь один пучок, в то же время у одного клубенька Sesbania grandiflora в возрасте одного года их удалось насчитать до 126. Довольно часто сосудистая система клубенька отделяется с внешней стороны от его коры слоем частично или полностью опробковевших клеток, получивших название клубеньковой эндодермы, соединенных с эндодермой корня. Клубеньковая эндодерма представляет собой внешний слой неинфицированной коровой паренхимы, расположенной между клубеньковой тканью и корневой корой.
У большей части видов растений клубеньки образуются по описанному типу. Следовательно, образование клубеньков — результат сложных явлений, начинающихся вне корня. Вслед за начальными фазами инфекции индуцируется образование клубенька, затем происходит распространение бактерий в зоне клубеньковой ткани и фиксация азота.
Все стадии развития клубеньковых бактерий, по данным чешского микробиолога В. Каша (1928), можно проследить на срезах клубеньков. Так, в верхней части клубенька, например, люцерны содержатся в основном мелкие делящиеся палочковидные клетки, в небольшом количестве молодые бактероиды, число которых возрастает постепенно по мере развития клубенька. В средней, окрашенной в розовый цвет части клубенька обнаруживаются преимущественно бактероидные клетки и реже мелкие палочковидные. В основании клубенька на ранних стадиях вегетации растения-хозяина бактероиды такие же, как и в средней его части, а к концу вегетации более раздутые и раньше дегенерирующие.
Сроки появления первых видимых клубеньков на корнях различных видов бобовых растений различны (М. В. Федоров, 1952). Появление их у большинства бобовых культур чаще всего происходит во время развития первых настоящих листьев. Так, образование первых клубеньков люцерны посевной наблюдается между 4-м и 5-м днями после прорастания, а на 7 — 8-й день этот процесс происходит у всех растений. Клубеньки у люцерны серповидной появляются через 10 дней.
В период функционирования клубеньки обычно плотные. Клубеньки, образованные активными культурами бактерий, в молодом возрасте имеют беловатую окраску. К моменту проявления оптимальной активности они становятся розовыми. Клубеньки, возникшие при инфекции неактивными культурами бактерий, зеленоватого тона. Нередко их структура практически не отличается от структуры клубеньков, образованных при участии активных штаммов клубеньковых бактерий, но они преждевременно разрушаются.
В некоторых случаях строение клубеньков, образуемых неактивными бактериями, отклоняется от нормы. Это выражается в дезорганизации клубеньковой ткани, утрачивающей обычно четко выраженную зональную дифференциацию.
Розовая окраска определяется наличием в клубеньках пигмента, по химическому составу близкого гемоглобину крови. В связи с этим пигмент называется леггемоглобином (легоглобином) — гемоглобином Leguminosae. Легоглобин содержится лишь в тех клетках клубеньков, в которых имеются бактероиды. Он локализован в пространстве между бактероидами и окружающей их мембраной.
Количество его колеблется от 1 до 3 мг на 1 г клубенька, в зависимости от вида бобового растения.
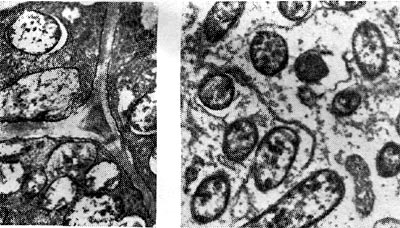
Рис. 162. Бактериодная ткань в период активного функционирования клубеньков клевера (слева увел. X 10 000) и вики посевной (справа — увел. X 6000).
У однолетних бобовых растений к концу вегетационного периода, когда заканчивается процесс азотфиксации, красный пигмент переходит в зеленый. Изменение цвета начинается у основания клубенька, позднее зеленеет его вершина. У многолетних бобовых растений позеленения клубеньков не происходит или оно наблюдается только у основания клубенька. У разных видов бобовых растений переход красного пигмента в зеленый происходит с разной степенью интенсивности и разной скоростью.
Клубеньки однолетних растений функционируют сравнительно недолго. У большинства бобовых культур некроз клубенька начинается в период цветения растения-хозяина и протекает обычно в направлении от центра к периферии клубенька. Один из первых признаков разрушения — образование слоя клеток с мощными стенками у основания клубенька. Этот слой клеток, расположенный перпендикулярно к главному сосуду корня, разъединяет его с клубеньком и задерживает обмен питательными веществами между растением-хозяином и тканями клубенька.
В клетках дегенерирующей ткани клубенька появляются многочисленные вакуоли, ядра теряют способность окрашиваться, часть клеток клубеньковых бактерий лизируется, часть мигрирует в окружающую среду в виде мелких кокковидных клеток-артроспор.
Процесс формирования артроспор в ткани лизирующегося клубенька показан на рисунках 163—165. Прекращают функционировать в этот период и инфекционные нити (рис. 166). Клетки хозяина утрачивают тургор и сжимаются теми соседними клетками, которым он еще свойствен.
Старые клубеньки темные, дряблые, мягкие. При надрезе из них выступает водянистая слизь. Процессу разрушения клубенька, начинающегося с опробковения клеток сосудистой системы, способствуют понижение фотосинтетической активности растения, сухость или чрезмерная влажность среды.
В разрушенном, ослизненном клубеньке обнаруживаются часто простейшие, грибы, бациллы и мелкие палочковидные клубеньковые бактерии.
Состояние растения-хозяина оказывает влияние на длительность функционирования клубенька. Так, по данным Ф. Ф. Юхимчука (1957), кастрируя или удаляя цветы люпина, можно продлить период его вегетации и вместе с тем время активной деятельности клубеньковых бактерий.
Клубеньки многолетних растений, в отличие от клубеньков однолетних, могут функционировать в течение многих лет. Так, например, карагана имеет многолетние клубеньки, в которых процесс старения клеток идет одновременно с образованием новых. У вистерии (глицинии китайской) также функционируют многолетние клубеньки, образуя на корнях хозяина шаровидные вздутия. К концу вегетационного периода бактероидная ткань многолетних клубеньков деградирует, но весь клубенек не отмирает. На следующий год он вновь начинает функционировать.
Дата добавления: 2020-11-18; просмотров: 634;
Поиск по сайту
Узнать еще
- I. Общая характеристика категории состояния как часть речи
- II. Лексико-грамматические разряды имен числительных. Их характеристика.
- II. Лексико-грамматические разряды местоимений. Их общая характеристика
- IV. ОБЩАЯ ХАРАКТЕРИСТИКА ВИРУСОВ
- U - образная характеристика
- XXIII. ОБЩАЯ ХАРАКТЕРИСТИКА
- А. Общая характеристика
- Абсолютные показатели экономичности и энергетическая характеристика оборудования

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
