Факторы, определяющие симбиотические взаимоотношения клубеньковых бактерий с бобовыми растениями
Для симбиоза, обеспечивающего хорошее развитие растений, необходим определенный комплекс условий среды. Если условия окружающей среды будут неблагоприятными, то, даже несмотря на высокую вирулентность, конкурентную способность и активность микросимбионта, эффективность симбиоза будет низкой.
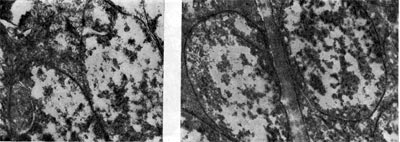
Рис. 163. Начальный период формирования артроспор в бактероидах клевера. Увел. X 30 000.
Для развития клубеньков оптимальная влажность 60—70% от полной влагоемкости почвы. Минимальная влажность почвы, при которой еще возможно развитие клубеньковых бактерий в почве, приблизительно равна 16% от полной влагоемкости. При влажности ниже этого предела клубеньковые бактерии обычно уже не размножаются, но тем не менее они не погибают и могут длительное время сохраняться в неактивном состоянии. Недостаток влаги приводит и к отмиранию уже сформировавшихся клубеньков.
Нередко в районах с недостаточным увлажнением многие бобовые растения развиваются, не образуя клубеньков.
Поскольку размножение клубеньковых бактерий в отсутствие влаги не происходит, в случае засушливой весны инокулированные (искусственно зараженные) семена необходимо вносить глубже в почву. Например, в Австралии семена с нанесенными на них клубеньковыми бактериями глубоко заделывают в почву. Интересно, что клубеньковые бактерии почв засушливого климата более стойко переносят засуху, чем бактерии почв влажного климата. В этом проявляется их экологическая приспособленность.
Избыточная влажность, как и ее недостаток, также неблагоприятна для симбиоза — из-за снижения степени аэрации в зоне корней ухудшается снабжение корневой системы растения кислородом. Недостаточная аэрация отрицательно влияет и на живущие в почве клубеньковые бактерии, которые, как известно, лучше размножаются при доступе кислорода. Тем не менее высокая аэрация в зоне корней приводит к тому, что кислород начинают связывать восстановители молекулярного азота, снижая степень азотфиксации клубеньков.
Важную роль во взаимоотношениях клубеньковых бактерий и бобовых растений играет температурный фактор. Температурные характеристики разных видов бобовых растений различны. Также и разные штаммы клубеньковых бактерий имеют свои определенные температурные оптимумы развития и активной фиксации азота. Следует отметить, что оптимальные температуры развития бобовых растений, образования клубеньков и азотфиксации не совпадают. Так, в природных условиях образование клубеньков может наблюдаться при температурах несколько выше О °С, азот-фиксация при таких условиях практически не происходит. Возможно, лишь арктические симбиозирующие бобовые растения связывают азот при очень низких температурах. Обычно же этот процесс происходит лишь при 10 °С и выше. Максимальная азотфиксация ряда бобовых растений наблюдается при 20—25 °С. Температура выше 30 °С отрицательно влияет на процесс азотонакопления.
Экологическая адаптация к температурному фактору у клубеньковых бактерий значительно меньше, чем у многих типичных сапрофитных форм. По мнению Е.Н. Мишустина (1970), это объясняется тем, что естественной средой обитания клубеньковых бактерий являются ткани растений, где температурные условия регулируются растением-хозяином.
Большое влияние на жизнедеятельность клубеньковых бактерий и образование клубеньков оказывает реакция почвы. Для разных видов и даже штаммов клубеньковых бактерий значение рН среды обитания несколько различно. Так, например, клубеньковые бактерии клевера более устойчивы к низким значениям р.Н, чем клубеньковые бактерии люцерны. Очевидно, здесь также сказывается адаптация мпкро-ерганизмов к среде обитания. Клевер растет на более кислых почвах, чем люцерна. Реакция почвы как экологический фактор оказывает влияние на активность и вирулентность клубеньковых бактерий. Наиболее активные штаммы, как правило, легче выделить из почв с нейтральными значениями рН. В кислых почвах чаще встречаются неактивные и слабовирулентные штаммы. Кислая среда (рН 4,0 — 4,5) оказывает непосредственное влияние и на растения, в частности нарушая синтетические процессы обмена веществ растений и нормальное развитие корневых волосков. В кислой среде у инокулированных растений резко сокращается срок функционирования бакте-роидной ткани, что ведет к снижению степени азотфиксации.
В кислых почвах, как отмечает А. В. Петербургский, в почвенный раствор переходят соли алюминия и марганца, неблагоприятно действующие на развитие корневой системы растений и процесс азотоусвоения, а также снижается содержание усвояемых форм фосфора, кальция, молибдена и углекислоты. Неблагоприятную реакцию почвы лучше всего устраняет известкование.
Размеры симбиотической азотфиксации определяются в значительной степени условиями питания растения-хозяина, а не клубеньковых бактерий. Клубеньковые бактерии как эндотрофные симбионты растений зависят в основном от растения при получении углеродсодер-жащих веществ и минеральных элементов питания.
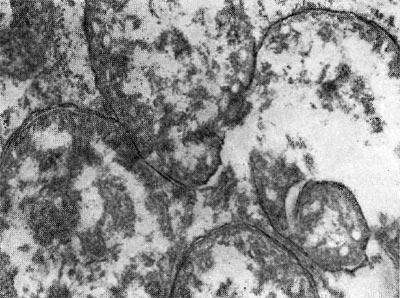
Рис. 164. Бактероид со сформировавшейся артроспорой клубеньковой бактерии клевера. Увел. X 35 000.
Для клубеньковых бактерий ткань хозяина представляет такую питательную среду, которая может удовлетворить даже самый требовательный штамм вследствие содержания в ткани всех типов питательных веществ. Тем не менее после внедрения клубеньковых бактерий в ткань растения-хозяина их развитие определяется не только внутренними процессами, но и в значительной степени зависит от действия внешних факторов, оказывающих влияние на весь ход инфекционного процесса. Содержание или отсутствие того или иного питательного вещества в окружающей среде может быть определяющим моментом для проявления симбиотической азотфиксации.
Степень обеспеченности бобовых растений доступными формами минеральных соединений азота определяет эффективность симбиоза. На основании многочисленных лабораторных и вегетационных опытов известно, что чем больше азотсодержащих соединений в окружающей среде, тем с большим трудом внедряются бактерии в корень.
Сельскохозяйственная практика требует однозначно решить задачу — целесообразнее удобрять бобовые культуры азотом или же правы те исследователи, которые утверждают, что минеральный азот подавляет симбиотическую азотфиксацию бобовых культур и поэтому экономически выгоднее такие растения азотом не удобрять. На кафедре агрономической и биологической химии Московской сельскохозяйственной академии им. К. А. Тимирязева были проведены опыты, результаты которых дали возможность получить картину поведения симбионтов в условиях вегетационных и полевых опытов при обеспеченности растений разными дозами азота в среде. Установлено, что повышение содержания растворимых азотсодержащих соединений в среде в полевых условиях при оптимальных условиях произрастания растений не препятствует их симбиозу с клубеньковыми бактериями. Снижение доли атмосферного азота, усваиваемого растениями при повышенной обеспеченности минеральным азотом, имеет только относительный характер. Абсолютное количество азота, усвоенного бактериями из атмосферы, практически не снижается, даже нередко увеличивается по сравнению с растениями, выращивающимися в присутствии клубеньковых бактерий, но без внесения в почву азота.
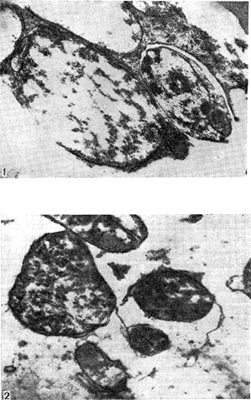
Рис. 165. Миграция артроспор в окружающую среду из клубеньков: 1 — клевера (увел. X 17 000), 2 — люцерны (увел. X 23 000).
Большое значение в активации усвоения азота бобовыми растениями имеет фосфорное питание. При низком содержании фосфора в среде проникновение бактерий в корень происходит, но клубеньки при этом не образуются. Бобовым растениям присущи некоторые особенности в обмене фосфорсодержащих соединений. Семена бобовых отличаются повышенным содержанием фосфора. Запасной фосфор при прорастании семян используется не так, как у других культур, — сравнительно равномерно для формирования всех органов, а в большей степени сосредоточиваясь в корнях. Поэтому в ранние сроки развития бобовые растения, в отличие от злаковых, в большей степени удовлетворяют свои потребности в фосфоре за счет семядолей, а не запасов почвы. Чем крупнее семена, тем меньше бобовые растения зависят от фосфора почвы. Однако при симбиотическом способе существования потребность бобовых растений в фосфоре выше, чем при автотроф-ном. Поэтому при недостатке фосфора в среде у инокулированных растений ухудшается снабжение растений азотом.
Бобовые растения, как известно, выносят с урожаем значительно больше калия, чем другие сельскохозяйственные культуры. Поэтому калийные и особенно фосфорно-калийные удобрения существенно повышают продуктивность азотфиксации бобовыми растениями.
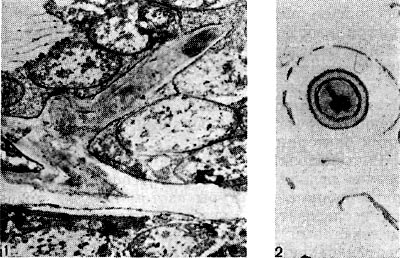
Рис. 166. Прекратившие функционировать интерцеллюлярная (1) и интрацеллюлярная (2) инфекционные нити в клубеньках клевера. Увел. X 10 000.
Положительное действие калия на образование клубеньков и интенсивность азотфиксации связано в значительной степени с физиологической ролью калия в углеводном обмене растения.
Кальций нужен не только для устранения излишней кислотности почвы. Он играет специфическую роль в развитии клубеньковых бактерий и обеспечении нормального симбиоза бактерий с растением-хозяином. Потребность клубеньковых бактерий в кальции частично может быть компенсирована стронцием. Интересно, что клубеньковые бактерии тропических культур, растущих на кислых латеритных почвах, не нуждаются в кальции. В этом опять проявляется экологическая адаптация клубеньковых бактерий, поскольку тропические почвы содержат очень небольшие количества кальция.
Для симбиотической азотфиксации необходимы также магний, сера и железо. При недостатке магния тормозится размножение клубеньковых бактерий, снижается их жизнедеятельность, подавляется симбиотическая азот-фиксация. Сера и железо оказывают также благоприятное влияние на образование клубеньков и процесс азотфиксации, в частности играя несомненную роль в синтезе леггемоглобина.
Из микроэлементов особо отметим роль молибдена и бора. При недостатке молибдена клубеньки плохо образуются, в них нарушается синтез свободных аминокислот и подавляется синтез леггемоглобина. Молибден вместе с другими элементами с переменной валентностью (Fe, Co, Си) служит посредником при переносе электронов в окислительно-восстановительных ферментных реакциях. При дефиците бора в клубеньках не формируются сосудистые пучки, и вследствие этого нарушается развитие бактероидной ткани.
На формирование клубеньков у бобовых растений большое влияние оказывает углеводный обмен растений, определяемый рядом факторов: фотосинтезом, наличием в среде углекислого газа, физиологическими особенностями растений. Улучшение углеводного питания благоприятно сказывается на инокуляционном процессе и азотонакоплении. С практической точки зрения большой интерес представляет использование соломы и соломистого свежего навоза для удобрения бобовых растений как источника углеводов. Но в первый год после внесения соломы в почву при ее разложении накапливаются токсические вещества. Следует отметить, что не все виды бобовых растений чувствительны к токсическим продуктам распада соломы; горох, например, не реагирует на них.
Определенное значение в симбиозе клубеньковых бактерий и бобовых растений имеют биологические факторы.
Большое внимание уделяется влиянию ризо-сферной микрофлоры на клубеньковые бактерии, которое может иметь как стимуляционный, так и антагонистический характер в зависимости от состава микроорганизмов ризосферы.
Много работ посвящено изучению фагов клубеньковых бактерий. Большинство фагов способны лидировать различные виды бактерий, некоторые специализированы лишь в отношении отдельных видов или даже штаммов клубеньковых бактерий. Фаги могут препятствовать внедрению бактерий в корень, вызывать лизис клеток в ткани клубенька. Фаги наносят большой ущерб, лизируя препараты клубеньковых бактерий на заводах, вырабатывающих нитрагин.
Среди различных видов насекомых, наносящих вред клубеньковым бактериям, особенно выделяется полосатый клубеньковый долгоносик, личинки которого разрушают клубеньки на корнях многих видов бобовых растений (главным образом однолетних). Широко распространен и щетинистый клубеньковый долгоносик.
Ранней весной самки клубеньковых долгоносиков откладывают от 10 до 100 яиц. Через 10—15 дней из яиц развиваются небольшие (до 5,5 мм), червеобразные, согнутые, белые, со светло-бурой головкой личинки, питающиеся преимущественно клубеньками и корневыми волосками. Только что вылупившиеся личинки проникают в клубенек и питаются его содержимым. Более взрослые личинки разрушают клубеньки снаружи. Одна личинка за 30—40 дней уничтожает 2—6 клубеньков. Особенно большой вред они наносят в сухую и жаркую погоду, когда развитие растений замедляется.
Клубеньки люцерны и некоторых других видов бобовых растений повреждает также большой люцерновый долгоносик.
Самки жука откладывают до 400 яиц, из которых развиваются безногие, дугообразные, желтовато-белые, с бурой головкой, покрытые бурыми щетинками личинки. Их длина 10— 14 мм. Цикл развития большого люцернового долгоносика протекает в течение двух лет.
Наличие нематод в корневой зоне различных видов бобовых растений отмечают многие исследователи. В прикорневой зоне гороха, например, обнаружено 47 видов нематод, среди них 25 паразитических.
На корнях молодых растений фасоли, люпина, клевера может паразитировать широко распространенная ростковая нематода. Самки этого вида, питающиеся корнями растений, откладывают яйца в ткани растения. Весь жизненный цикл развивающейся из яиц нематоды протекает обычно внутри тканей.
В степных районах на корнях люцерны, клевера и сои обнаружена степная нематода. Самки перед откладкой яиц проникают в корень, куда откладывают от 12 до 20 яиц. В корнях личинки проходят три личиночные стадии развития, нарушая функции корня и клубеньков.
Дата добавления: 2020-11-18; просмотров: 715;
Поиск по сайту
Узнать еще
- Анализ объекта оценки с позиции отражающей взаимоотношения компонентов собственности
- Антропогенные факторы, влияющие на биоразнообразие
- Биология аэробных спорообразующих бактерий
- Биотические взаимоотношения в биосфере, их закономерности. Биоразнообразие и его значение. (Модель Н.Ф. Реймерса “Волчок жизни”)
- Биотические взаимоотношения между популяциями и организмами
- Биохимические механизмы резистентности бактерий к антибиотикам
- БОБОВЫМИ И МАКАРОННЫМИ ИЗДЕЛИЯМИ
- В теории двойственности важное значение имеют две теоремы, определяющие связь между решениями прямой и двойственной задач.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
