Физиология спорообразования у анаэробов
Причины, вызывающие переход бактерий от вегетативного развития к спорообразованию, длительное время были неизвестны микробиологам. Впервые в нашей стране А. Я. Мантеифель, Н. Д. Иерусалимский провели исследования, вскрывшие факторы, влияющие на образование спор сахаролитическими анаэробами. Оказалось, что спорообразование вызывается исчезновением из среды источников азота, а также накоплением продуктов обмена (например, бутилового спирта). Но чтобы приобрести способность к споруляции, бактерии должны развиваться при нормальных условиях на полноценной питательной среде, содержащей набор аминокислот и факторов роста. Кислотность среды, оптимальная для вегетативного роста, является наиболее благоприятной и для спорообразования (рН 4,5—5,5 для Cl. acetobutylicum и ~7—7,2 для протеолитических анаэробов). Аэрация останавливает споруляцию. Эти данные свидетельствуют о том, что спорообразование является нормальной стадией развития у этих бактерий.
У анаэробов в отличие от аэробных видов исчерпание в среде источников углерода не влияет на процесс спорообразования. Даже добавление к среде глюкозы (5 г на литр) не сказывается на процессе спорообразования. Если культура выращена на полноценной питательной среде и приступила к спорообразованию, то дальнейший перевод клеток в условия полного голодания (например, перенос в водопроводную воду) не только не подавляет, но даже стимулирует споруляцию у некоторых сахаролитических анаэробов (Cl. saccharobutyricum), т. е. для спорообразования не требуется в этом случае поступления питательных веществ из внешней среды (экзогенных источников питания), а для завершения конечных этапов формирования спор используются внутриклеточные запасы. Такой обмен веществ называется эндогенным (или эндотрофным) метаболизмом. Действительно, установлено, например, что белок спор образуется за счет азота белка вегетативных клеток. Однако у протеолитических анаэробов споруля-ция не поддерживается в среде, полностью лишенной органических питательных веществ. Так, для спорообразования Cl. botulinum требуются аминокислоты — аланин и аргинин, которые используются в качестве источников энергии.
>Генетический контроль спорообразования.
Процесс «поглощения» материнской клеткой от сеченной на полюсе части цитоплазмы с ядерным элементом приводит к образованию новой клетки с двумя мембранами (проспора). Возникает особая ассоциация клеток — двуклеточ-ный организм в общей оболочке, причем одна из клеток — проспора осуществляет свою жизнедеятельность внутри цитоплазмы другой, материнской клетки. Возникает крайне интересная ситуация, где сталкивается деятельность двух генетических центров — ядра материнской клетки и ядра споры. Многие стороны в их взаимоотношениях остаются еще неясными. Но из цитологических, физиологических и генетических наблюдений можно сделать вывод, что эти два ядра «разделяют» между собой обязанности, связанные с контролем над процессами формирования споровых структур.
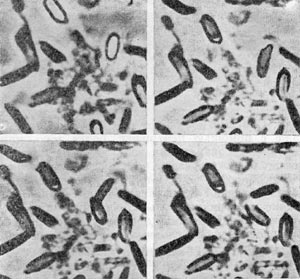
Рис. 63. Стадии прорастания спор Cl. sporopenitum. Увел. X 2100.
Так, синтез новых белковых веществ, споровых оболочек, кортекса и экзоспориума осуществляется при активном контроле генома материнской клетки. При этом роль материнской клетки сводится к тому, чтобы как-то отделить от себя дочернюю клетку, оказавшуюся внутри ее цитоплазмы. Но так как удалить проспору за пределы клеточных границ через цитоплазматическую мембрану и клеточную стенку невозможно, то материнская клетка поступает с дочерней по-иному — она изолирует ее от себя путем инцистирования, окружая проспору мощными слоями белковой оболочки и переводя ее в метаболически неактивное (анабиотическое) состояние. Но это все равно не спасает материнскую клетку от гибели. Хотя она и продолжает еще длительное время активно функционировать, даже иногда способна образовать еще одну спору, но все же она не в состоянии вернуться к вегетативному развитию. В результате нарушения обмена веществ в конце концов наступает лизис (разрушение) вегетативной материнской клетки и освобождение спор.
Участие в спорогенезе большого количества ферментов и антигенов, включение новых путей биосинтеза и появление новых специфических клеточных структур показывает, что число генов, контролирующих спорогенез, достаточно велико (по всей вероятности, их больше 100). Некоторые из этих генов функционируют и в период вегетативной стадии развития. До сих пор остается неясным механизм функционирования спорового генома (набора генов, ответственных за спорообразование). Установлено, что споры получают весь свой ядерный материал (ДНК) в готовом виде от материнских клеток. При этом количество ДНК на спору постоянно для данного вида и не зависит от среды выращивания бактерий. Таким образом, этот показатель, вероятно, может служить характеристикой вида и использоваться при систематике спорообразую-щих бактерий.
Прорастание спор
Споры, перенесенные в свежую питательную среду, начинают прорастать. Сначала они набухают, темнеют, затем через образовавшееся отверстие в оболочке споры молодая клетка выходит в среду. При этом слой кортекса разрушается, а споровая оболочка вместе с отростками (если таковые имеются) сбрасывается. У анаэробов проследить за прорастанием, наблюдая за одной и той же спорой, удается только в редких случаях. На рисунке 63 представлена серия фотографий, иллюстрирующих различные стадии прорастания — от потемнения сердцевины спор до выхода молодой вегетативной клетки из споровой оболочки. На последней стадии выхода вегетативная клетка как бы отстреливается. Отверстие в оболочке споры образуется не строго на полюсе споры, а несколько сбоку, и молодая вегетативная клетка при выходе расположена под углом к длинной оси споры. У других анаэробов процесс прорастания может выглядеть иначе.
Особенности прорастания Cl. pasteurianum С. Н. Виноградскии (1902) использовал для дифференциации этого вида от других спорообразующих анаэробов. Наконец, три вида клостридиев — Cl. pectinovorum, Cl. bu-tyricum и Cl. tetani — отличаются тем, что прорастание их спор происходит внутри спорангия. Клеточная стенка (или часть ее) у этих видов не лизируется, а остается на зрелых спорах, облегая их в виде чехла. Но этот чехол не идентичен по происхождению и строению с экзоспориумом, описанным выше.
Дата добавления: 2020-11-18; просмотров: 730;
Поиск по сайту
Узнать еще
- АНАТОМИЯ И ФИЗИОЛОГИЯ ОРГАНОВ ДЫХАНИЯ.
- Анатомия и физиология поджелудочной железы
- Анатомия и физиология почек
- Анатомия и физиология сердца
- АНАТОМИЯ И ФИЗИОЛОГИЯ ЧЕЛОВЕКА КАК НАУКА
- Анатомия и физиология щитовидной железы
- Анатомия и физиология — науки о строении и функциях организма.
- АНАТОМИЯ, ГИСТОПАТОЛОГИЯ, ФИЗИОЛОГИЯ КОЖИ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
