Глава 14 БИОДИНАМИКА (БИОМЕХАНИКА) ДВИГАТЕЛЬНЫХ КАЧЕСТВ
Каждый человек владеет определенными двигательными навыками, например, может поднять определенный вес, пробежать или прыгнуть и т. п., но возможности у всех различны. Это связано и с возрастом, и наследственностью и, главное, с тренированностью. Двигательные качества отличаются друг от друга по форме и по затраченной энергии. Двигательные качества — это отдельные стороны моторики человека. Они проявляются в одинаковой форме движений и энергетического обеспечения и имеют аналогичные физиологические механизмы.
Поэтому методики совершенствования (тренировки) тех или иных качеств имеют общие черты независимо от конкретного вида движения. Например, выносливость марафонца во многом сходна с выносливостью лыжника-гонщика, велогонщика, конькобежца и т. п. Сила (F), скорость (V) и длительность (продолжительность) (t) движения находятся в определенных соотношениях друг с другом. Это соотношение различно в разных видах деятельности (в разных видах спорта).
При сокращении мышцы развивают большие усилия, которые зависят от поперечного сечения, начальной длины волокон и ряда других факторов. Сила мышцы на 1 см2 ее поперечного сечения называется абсолютной мышечной силой. Для человека она равна от 50 до 100 Н.
Сила и мощность одних и тех же мышц зависят от ряда физиологических условий: возраста, пола, тренировки, температуры воздуха, исходного положения при выполнении упражнений, биоритмов и т. д..
Внешнее проявление сократительной активности мышцы (пучка волокон или волокна) состоит в том, что при ее фиксированной
длине развивается усилие, а при фиксированной нагрузке происходит укорочение. Эксперимент с мышцами проводится в двух режимах: изометрическом, когда длина мышцы фиксирована и изотоническом, когда мышца имеет возможность укорачиваться при постоянной нагрузке (рис. 14.1). На рисунке видно, что изометрическое усилие развивается очень быстро и достигает своей максимальной величины примерно через 170 мс после возбуждения. Начиная с 200 мс оно снова уменьшается с возрастающей скоростью. Интересно отметить, что даже через 900 мс в мышце еще сохраняется некоторое напряжение, что может быть обусловлено только активными физическими и химическими процессами.
Изотоническое одиночное сокращение существенно отличается от изометрического. Укорочение в процессе изотонического одиночного сокращения начинается только тогда, когда в мышце развивается достаточное усилие, равное по величине внешнему. В результате одиночное сокращение начинается тем позднее, чем больше нагрузка. Укорочение вначале почти линейно зависит от времени и достигает максимальных значений тем раньше, чем больше нагрузка. Затем наступает расслабление мышц с возрастающей скоростью, причем, так же как и укорочение, оно завершается тем раньше, чем больше груз. Если сделать нагрузку равной тому полному изометрическому усилию, которое мышца способна развить, то никакого внешнего укорочения не произойдет. При нулевой нагрузке скорость укорочения, очевидно, должна быть максимальной. Соотношение между нагрузкой и установившейся скоростью укорочения показано на рис. 14.2.
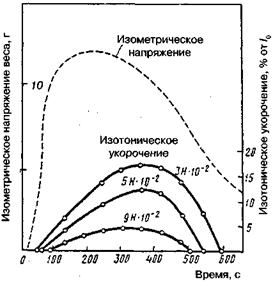
Рис.14.1. Изометрическое и изотоническое одиночное сокращение. Портняжная мышца лягушки при 0°С (по В. Jewell, D. Wilkie, 1960)
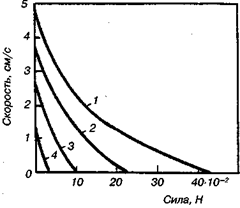
Рис. 14.2.Зависимость скорости от нагрузки на различных стадиях изотонического одиночного сокращения портняжной мышцы лягушки при 0°С: 1 — фаза развития напряжения, 2—4 — фазы расслабления (0,46; 0,64; 0,83 с); напряжение составляет 0,6; 0,3 и 0,08 максимального

V== b(F0-F)·(F + a)
или
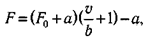
где V — скорость укорочения; F — сила (нагрузка); F0 — максимальная изометрическая сила, которую может развить мышца; b — константа, имеющая размерность силы. Максимальная скорость, соответствующая условно F = О, из уравнения Хилла равна 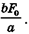 При раздражении мышцы серией импульсов, следующих с постоянной частотой, второй и последующие импульсы будут оказывать разное действие в зависимости и от того, на какой участок кривой «сила — время» они попадут. Например, для портняжной мышцы лягушки при 0°С (рефракторный период потенциала действия около 10 мс) второй импульс, отстоящий от первого на 5 мс, не вызывает никакой дополнительной механической реакции. При частоте возбуждения, равной 2 Гц, импульсы будут поступать в момент, когда фаза расслабления завершится на 2/3. Мышца отреагирует следующим одиночным сокращением, которое, в свою очередь, прервется, не завершившись, новым импульсом и т. д. В результате получается кривая, каждый максимум которой соответствует одиночному импульсу. При соответствующем выборе частоты раздражения одиночные сокращения буду иметь все большую тенденцию к слиянию. У портняжной мышцы при 0°С наступает полное слияние — тетанус — при частоте примерно 15 Гц. Эффект слияния проявляется в том, что активное усилие увеличивается в 1,2—1,8 раза по сравнению с максимальным усилием одиночного сокращения. Следует отметить, что при одиночном импульсе активность целой мышцы не успевает достичь своего тетанического максимума, так как полное растяжение системы последовательных упругих элементов требует времени, которое больше длительности одиночного сокращения.
При раздражении мышцы серией импульсов, следующих с постоянной частотой, второй и последующие импульсы будут оказывать разное действие в зависимости и от того, на какой участок кривой «сила — время» они попадут. Например, для портняжной мышцы лягушки при 0°С (рефракторный период потенциала действия около 10 мс) второй импульс, отстоящий от первого на 5 мс, не вызывает никакой дополнительной механической реакции. При частоте возбуждения, равной 2 Гц, импульсы будут поступать в момент, когда фаза расслабления завершится на 2/3. Мышца отреагирует следующим одиночным сокращением, которое, в свою очередь, прервется, не завершившись, новым импульсом и т. д. В результате получается кривая, каждый максимум которой соответствует одиночному импульсу. При соответствующем выборе частоты раздражения одиночные сокращения буду иметь все большую тенденцию к слиянию. У портняжной мышцы при 0°С наступает полное слияние — тетанус — при частоте примерно 15 Гц. Эффект слияния проявляется в том, что активное усилие увеличивается в 1,2—1,8 раза по сравнению с максимальным усилием одиночного сокращения. Следует отметить, что при одиночном импульсе активность целой мышцы не успевает достичь своего тетанического максимума, так как полное растяжение системы последовательных упругих элементов требует времени, которое больше длительности одиночного сокращения.
В описанных экспериментах (исследованиях) изотоническое укорочение или изометрическое усилие измерялось на мышцах, длина которых была близка к длине расслабленной мышцы или несколько превосходила ее.
Исходя из термодинамики, химии и механики процесса сокращения и связи их с распространением возбуждения, П.И. Усиком, С.А. Ригерера (1973) сделаны предпосылки модели: а) мышца производит работу за счет прямого преобразования энергии, освобождающейся при механохимических реакциях; б) механохимические реакции протекают в большом числе малых, но конечных областей, распределенных по всему объему мышцы; в) источники исходных химических реагентов также распределены по всему объему мышцы; г) мышечная ткань анизотропна и обладает упругими и вязкими свойствами, причем вязкость обусловлена преимущественно миофибриллами, а упругость — соединительной тканью и другими структурами.
Дата добавления: 2020-11-18; просмотров: 608;
Поиск по сайту
Узнать еще
- F82 Специфическое расстройство развития двигательных функций
- H. Разработка мер по повышению качества работы органа здравоохранения
- IV. Степени качества наречий
- Mетодические критерии качества измерений в социальных науках
- VII. ОБСЛЕДОВАНИЕ ПОМЕТОВ И КОНТРОЛЬ КАЧЕСТВА ИХ ВЫРАЩИВАНИЯ
- Індивідуальний стиль 1 глава
- Індивідуальний стиль 2 глава
- Індивідуальний стиль 3 глава

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
