ЧЕЛНОЧНЫЕ МЕХАНИЗМЫ
Цитозольный НАДH (реакция 6 гликолиза) не может передавать водород на дыхательную цепь, потому что митохондриальная мембрана для него непроницаема. Перенос водорода через мембрану происходит с помощью специальных систем, называемых "челночными". Водород транспортируется через мембрану при участии пар субстратов, связанных соответствующими дегидрогеназами, т.е. с обеих сторон митохондриальной мембраны находится специфическая дегидрогеназа.
Глицеролфосфатная челночная системаработает в клетках белых мышц, печени, мозге.
Водород от НАДH в цитозоле передаётся на диоксиацетонфосфат глицерол-3-фосфатдегидрогеназой (НАД-зависимый фермент). Образовавшийся глицерол-3-фосфат окисляется ферментом внутренней мембраны митохондрий - глицерол-3-фосфатдегидрогеназой (ФАД-зависимый фермент). Затем протоны и электроны с ФАДH2 переходят на убихинон и далее по дыхательной цепи.
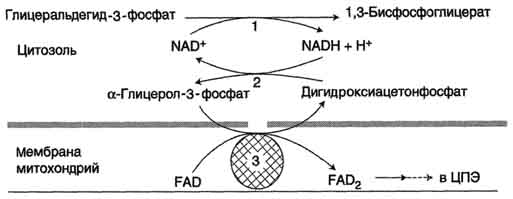
1 - глицеральдегид-3-фосфатдегидрогеназа;
2 - глицерол-3-фосфатдегидрогеназа (цитозольный фермент);
3 - глицерол-3-фосфатдегидрогеназа (митохондриальныи фермент).
Малат-аспартатная челночная система в которой участвуют малат, цитозольная и митоховдриальная малат-дегидрогеназы, является более универсальной, и работает в сердечной мышце, печени, почках.
В цитоплазме НАДH восстанавливает оксалоацетат в малат (реакция 1), который при участии переносчика проходит в митохондрии, где окисляется в оксалоацетат НАД-зависимой малатдегидрогеназой (реакция 2). Восстановленный НАДН отдаёт водород в митохондриальную ЦПЭ.
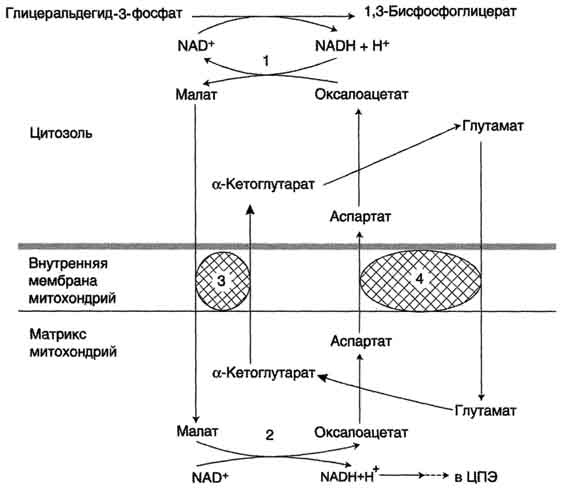
1,2 - окислительно-восстановительные реакции, обеспечивающие транспорт водорода из цитозоля в митохондрии на ЦПЭ;
3,4 - транслоказы, обеспечивающие транспорт a-кетоглутарата, аспартата и глутамата и через мембрану митохондрий.
Образованный из малата оксалоацетат выйти из митохондрий в цитозоль не может: мембрана митохондрий для него непроницаема. Поэтому оксалоацетат превращается в аспартат, который транспортируется в цитозоль, где снова превращается в оксалоацетат.
Обе челночные системы существенно отличаются по количеству синтезированного АТФ. В первой системе – 2АТФ (водород вводится в ЦПЭ на уровне убихинона). Вторая система энергетически более эффективна – 3АТФ (водород поступает в ЦПЭ с митохондриальным НАД+).
ЦИКЛ КОРИ
Цикл Кори (глюкозо-лактатный цикл) открыла чешская ученая, лауреат Нобелевской премии Тереза Кори. Он представляет собой биохимический транспорт лактата из мышц в печень и дальнейший синтез глюкозы из лактата, катализируемый ферментами глюконеогенеза:
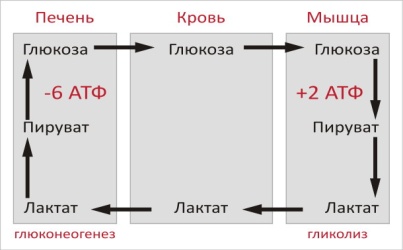
При интенсивной мышечной работе и в условиях отсутствия или недостаточного числа митохондрий (например, в эритроцитах) глюкоза подвергается анаэробному гликолизу с образованием лактата. При накоплении лактата в мышцах возникает лактат-ацидоз, раздражаются чувствительные нервные окончания, что вызывает боль в мышцах.
Лактат переносится кровью в печень и превращается в пируват, а затем в глюкозу (глюконеогенез), которая с током крови может возвращаться в работающую мышцу.
Направление лактатдегидрогеназной реакции в работающих мышцах и печени обусловлено отношением концентраций восстановленной и окисленной форм НАД+: отношение НАД+/НАДН в сокращающейся мышце больше, чем в печени.
СПИРТОВОЕ БРОЖЕНИЕ
Спиртовое брожение осуществляется дрожжеподобными организмами, а также некоторыми плесневыми грибками:
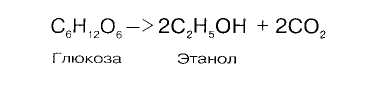
Механизм реакции близок к гликолизу. Расхождение начинается после этапа образования пирувата. При гликолизе пируват при участии фермента ЛДГ и кофермента НАДН восстанавливается в лактат. При спиртовом брожении пируват подвергается декарбоксилированию, в результате образуется ацетальдегид, а затем при восстановлении его - этанол:
При молочнокислом брожении ПВК не декарбоксилируется, а, как и при гликолизе в животных тканях, восстанавливается при участии ЛДГ за счет водорода НАДН.
Дата добавления: 2016-07-27; просмотров: 4553;
Поиск по сайту
Узнать еще
- III. Механизмы регуляции количества ферментов
- Безусловнорефлекторные, условнорефлекторные, гуморальные механизмы регуляции половых функций.
- Бесчелночные ткацкие станки
- Биологические и социальные аспекты старения. Генетические, молекулярные, клеточные, системные механизмы старения. Проблема долголетия.
- Биофизические механизмы транспорта вещества через биомембраны.
- Виды и механизмы коррозии стальных трубопроводов. Методы защиты МН от коррозии
- Виды, механизмы и цели познавательной деятельности
- Винтовые механизмы.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
