Ретикулярная формация ствола мозга
Ретикулярная формация – это система, которая объединяет анатомически и цитоархитектонически дискретные области ствола мозга, образованная скоплениями интернейронов разных ретикулярных типов. Морфологическими исследованиями в стволе мозга обнаружено несколько десятков обособленных групп клеток, обозначаемых как ядра и подъядра ретикулярной формации. Более удобным оказался принцип деления ретикулярной формации на медиальную и латеральную. На уровне перекреста пирамид (продолговатый мозг), где весь объем ретикулярной формации представлен латеральной областью, выделяют дорсальное и вентральное ретикулярные ядра.
Медиальная область ретикулярной формации начинается на уровне ядра подъязычного нерва парамедианным ретикулярным ядром, которое затем переходит в ретикулярное гигантоклеточное ядро продолговатого мозга. В мосту медиальная область представлена каудальным и оральным ретикулярными ядрами моста. Наиболее отчетливо различия между медиальной и латеральными областями обнаруживаются в продолговатом мозге и мосту, но они менее выражены в среднем. Вдоль средней линии на уровне продолговатого мозга и моста обнаружены скопления нервных клеток, обозначаемые как ядра шва. В среднем мозге к ним примыкают ядра околоводопроводного серого вещества. Эти ядра по ряду особенностей сходны с ретикулярной формацией. Наиболее общим типом нейронов, обнаруженных в ретикулярной формации, являются клетки с дихотомизированными аксонами, которые образуют восходящую и нисходящую ветви. В ретикулярной формации не обнаружены клетки II типа Гольджи, имеющие короткий аксон, профузно разделяющийся вблизи сомы на множество ветвей. Дендриты ретикулярных нейронов длинные и ветвящиеся, характерные для нейронов изодендритного типа; по количеству дендритов в медиальной области ретикулярной формации выделяют олигодендритные (1-2 дендрита) и полидендритные (до 6 дендритов) нейроны. Дендритные ветви ретикулярных нейронов ориентированы преимущественно в плоскости, перпендикулярной продольной оси мозга, хотя есть и некоторые отличия в распределении дендритов между нейронами медиальной и латеральной областей ретикулярной формации.
Ретикулярная формация тесно связана с различными структурами центральной нервной системы. От больших и малых нейронов медиальной ретикулярной формации к спинному мозгу направляются два наиболее развитых пути – ретикуло-спинальные тракты. От каудального и частично орального ретикулярных ядер моста начинается неперекрещивающийся понтинный, или медиальный, ретикуло-спинальный тракт, а от гиганто-
клеточного ядра продолговатого мозга берет начало билатеральный медуллярный, или латеральный, ретикуло-спинальный тракт. Волокна этих трактов достигают наиболее каудальных сегментов спинного мозга. Восходящие пути возникают из трех областей медиальной ретикулярной формации: из каудальной половины продолговатого мозга, из области на границе между продолговатым мозгом и мостом и из мезэнцефалической ретикулярной формации. Большинство восходящих волокон из продолговатого мозга и моста оканчиваются в неспецифических ядрах таламуса; окончания волокон мезэнцефалической ретикулярной формации обнаруживаются в гипоталамическом ядре преоптической области, септальных ядрах, таламическом ретикулярном ядре, медиальном и латеральном коленчатом телах таламуса, вентробазальном комплексе. Рассеянные волокна, принадлежащие понто-мезэнцефалической ретикулярной формации, обнаружены во многих областях переднего мозга, в том числе во фронтальной коре.
Важной особенностью ретикулярной формации является существование в ней большого количества ретикулярных нейронов, посылающих одновременно крупные коллатерали аксонов в спинной мозг и таламус. Эфферентные связи ретикулярной формации с мозжечком осуществляются через латеральное ретикулярное ядро и через парамедианное ретикулярное ядро покрышки моста. Ретикулярная формация получает большое количество афферентных волокон из различных источников. Наиболее интенсивны спиноретикулярные волокна от интернейронов V и VII пластин дорсальных рогов спинного мозга: 1) дорсальный пучок, оканчивающийся на нейронах медиальной области ретикулярной формации и моста, в латеральном ретикулярном ядре; 2) вентролатеральный пучок, содержащий спиноцеребеллярные, спинотектальные и спиноталамические пути. Окончания вентрального пучка обнаружены на нейронах ретикулярной формации среднего мозга. К нейронам ретикулярной формации поступает афферентация от сенсорных ядер тройничного, блуждающего, языкоглоточного, вестибулярного и слухового нервов. С корой ретикулярная формация связана через кортико-ретикулярные волокна из соматосенсорной коры. Ретикулярная формация получает церебелло-ретикулярные волокна от медиальных (фастигиальных) ядер мозжечка, которые оканчиваются в медиальной области ретикулярной формации продолговатого мозга и моста, а также в латеральном ретикулярном ядре. В ретикулярной формации среднего мозга находится красное ядро.
Красное ядромлекопитающих представлено скоплением нейронов в покрышке среднего мозга, локализующихся вентральнее глазодвигательных ядер и дорсального перекреста покрышки подбугорной области промежуточного мозга. Данная структура состоит из двух частей: ростральной – мелкоклеточной и каудальной – крупноклеточной. В составе красного ядра имеется три типа нейронов: большие, средние и мелкие. Медиатором тормозных интернейронов красного ядра является ГАМК. ГАМКергические нейроны преобладают в ростральной части ядра. В качестве кандидата в медиаторы в красном ядре выступает глутамат. Основным медиатором в красном ядре предположительно является ацетилхолин. Красные ядра имеют достаточно широкие связи, как афферентные, так и эфферентные, с различными структурами ЦНС. Переключение импульсации, вероятно, осуществляется на интернейронах ядра. Интернейроны красного ядра участвуют в полисинаптических реакциях эфферентных нейронов. Известно, что красное ядро является важнейшей станцией, объединяющей структуры стриопаллидарной системы, мозжечка и других экстрапирамидных образований.
Нисходящие влияния красного ядра осуществляются двумя путями: прямым, через рубро-спинальный тракт, и через ретикулярную формацию.
Аксоны крупноклеточной части красных ядер образуют мощные пучки волокон, которые сразу переходят на противоположную сторону, образуя форелевский переход, а затем спускаются в спинной мозг в виде рубро-спинального тракта.
Рубро-ретикулярные волокна образуют два пути:
1. Контрлатеральный рубро-бульбарный (является частью рубро-спинального тракта). Его волокна оканчиваются на нейронах латерального ретикулярного и мелкоклеточного ретикулярного ядер.
2. Рубро-ретикуло-оливарный тракт.Он берет начало от нейронов мелкоклеточной части красного ядра. Волокна этого тракта идут ипсилатерально к нейронам гигантоклеточного ретикулярного ядра, а также к оральному и каудальному ретикулярным ядрам моста.
Показаны также связи коллатералей рубро-спинальных нейронов с ядром лицевого нерва, главным тройничным ядром, нижним вестибулярным ядром, каудальным и главным клиновидными ядрами, нежным ядром продолговатого мозга.
Среди других эфферентных выходов из красных ядер выделяют рубро-оливарный тракт из мелкоклеточной части красного ядра в дорсальную пластину ипсилатеральной основной оливы, рубро-мозжечковый тракт к ядрам и коре мозжечка. Описан также неперекрещенный рубро-талами-
ческий эфферентный выход из красного ядра в вентролатеральное ядро таламуса, парафасцикулярный комплекс, срединный центр и ретикулярное таламическое ядро. Обнаружены также неперекрещенные рубронигральные связи, представленные в основном коллатералями рубро-спинальных нейронов.
Описаны афферентные проекции в красное ядро из мозжечка. Они являются контрлатеральными. Терминали волокон переднего промежуточного ядра мозжечка оканчиваются в крупноклеточной части красного ядра, а заднего промежуточного ядра мозжечка – в медиальном отделе мелкоклеточной части. Терминали большинства волокон зубчатого и латерального ядер мозжечка оканчиваются в мелкоклеточной части красного ядра, а у крысы и кошки встречаются и в крупноклеточной. Установлено, что мозжечково-рубральные волокна представлены не самостоятельными аксонами, а коллатералями мозжечково-таламических волокон. Так, в крупноклеточную часть красного ядра идут коллатерали аксонов промежуточного ядра, а в мелкоклеточную – коллатерали аксонов нейронов латерального ядра. Другим важным афферентным входом в красные ядра являются проекции из коры больших полушарий головного мозга. Показано, что у подавляющего числа млекопитающих кортико-рубральные волокна, идущие из сенсомоторных областей коры в красные ядра, организованы соматотопически. Причем наблюдается высокая степень конвергенции кортикальных нейронов на одиночных нейронах мелкоклеточной части красного ядра. Наряду с проекциями из сенсомоторных областей коры описаны проекции в красные ядра и из других областей коры мозга, в частности, из ассоциативной теменной коры мозга. Рубро-спинальные нейроны всех млекопитающих находятся под контролем коры головного мозга. Спинорубральные проекции также организованы по соматотопическому принципу: в дорсомедиальную область красного ядра проецируются афференты от ростральных частей тела и передних конечностей, а в вентральную область – афференты от каудальных частей тела и задних конечностей. Соматотопическое представительство спинорубральной проекции совпадает с таковым рубро-спинальной, мозжечково-рубральной и кортико-рубральной. Обнаружены также прямые проекции в красные ядра из бледного шара, энтопендункулярного ядра и черной субстанции. Установлены также прямые связи между красными ядрами и центральным серым веществом, головкой хвостатого ядра, центральным миндалевидным ядром, субталамическим, вестибулярными ядрами и голубым пятном.
Между кортико-спинальным трактом (пирамидная система) и рубро-спинальным трактом (экстрапирамидная система) существуют взаимные связи. Во-первых, данные системы имеют общие афференты: первичное моторное поле (поле 4), поле 6 коры больших полушарий. Во-вторых, терминали обеих систем перекрываются в одних и тех же слоях спинного мозга (вставочные нейроны V и VI пластины по Рекседу) и могут иметь синаптические контакты на одних и тех же нейронах. Имея обширные связи со структурами ЦНС, красное ядро участвует в организации различных типов движения через регуляцию мышечного тонуса, через изменение активности проприоспинальных систем, передающих нисходящие сигналы к мотонейронам. Поскольку интернейроны спинного мозга вовлечены в различные рефлекторные пути, красное ядро участвует в формировании моторного ответа посредством выборочного влияния на разные элементы выполняемого движения.
Медиаторные системы ретикулярной формации ствола мозга.
В стволе мозга существует обширная проприоретикулярная система, нейроны которой объединяют между собой ядра разных уровней ретикулярной формации, а также моторные и сенсорные ядра всех черепно-мозговых нервов. В ретикулярной формации находится система, содержащая специфические амины – серотонин, дофамин и норадреналин. Серотонинсодержащие нейроны локализуются у средней линии на всем протяжении продолговатого мозга, моста и среднего мозга. Серотонинергическая система образована ядрами шва. Стимуляция ядер шва оказывает тормозное действие на многие функции мозга (полисинаптические бульбарные и спинальные рефлексы, ретикулярные нейроны, нейроны коры). Показано, что серотонинергическая система входит в мощную и относительно специфическую систему нейронов ствола мозга, предназначенную для угнетения боли, которая имеет эфферентные пути к ноцицептивным структурам спинного мозга. Катехоламины содержатся в ядрах, расположенных ближе к средней линии в ростральной части среднего мозга. Катехоламинергическая система делится на норадренергическую и дофаминергическую. Нейроны, содержащие норадреналин, содержатся в латеральной части покрышки моста, а также в межуточном мозге. Существенное место в катехоламинергической системе занимает голубое пятно,которое локализуется в мосте и проецируется почти во все области мозга (кору больших полушарий, мозжечок, гипоталамус, различные структуры ствола мозга, спинной мозг). Нейроны голубого пятна являются донорами норадренергической системы. Полагают, что роль системы нейронов голубого пятна заключается в интеграции и передаче информации об уже наступивших изменениях состояния различных систем организма. Норадренергическую систему вообще считают интегратором поведенческого возбуждения и автономных функций.
В настоящее время голубое пятно интенсивно и всесторонне изучается. Считается, что голубое пятно является основной структурой норадренергической системы как в эмбриональном периоде, так и у взрослых млекопитающих. Микробиохимический и гистофлуоресцентный анализы показали, что голубое пятно представляет собой наибольшее скопление норадренергических нейронов в стволе мозга. Концентрация норадреналина в голубом пятне наивысшая по сравнению с другими исследованными ядрами мозга (900 нг/г белка). В настоящее время, кроме голубого пятна, в стволе мозга обнаружено еще несколько норадренергических зон – зоны А1, А2, А5. Эти образования изучены еще недостаточно. В структурах голубого пятна наряду с норадреналином содержится ряд других биологически активных веществ. В мелких гранулярных пузырьках (аппарат Гольджи) обнаружены серотонин, вещество Р, нейропептид галанин, имеющий широкий спектр действия, L-глутамат, аскорбиновая кислота, в телах нейронах голубого пятна найден дофамин.
В последние годы все большее внимание уделяется анализу рецепторов в голубом пятне. На соме и около сомы клеток голубого пятна выявлены α2-адренорецепторы и обнаружены также α1-адренорецепторы. В голубом пятне имеются глутаматрецепторы, выявлены ГАМК-рецепторы, опиатные рецепторы, обнаружены пуринергические рецепторы, галанинрецепторы. Установлено, что нейроны голубого пятна, способны функционировать как СО2/рН-хемосенсоры. Кроме α1- и α2-адренорецепторов в голубом пятне есть и β-адренорецепторы. Разница между этими рецепторами заключается в том, что α-адренорецепторы в качестве вторичных посредников используют инозилтрифосфат (ИТФ), диацилглицин (ДАГ) и ионы Са2+, а β-адренорецепторы соединены с ферментом аденилциклазой, продуцирующей циклический аденозинмонофосфат (цАМФ). α2-адренорецеп-
торы подразделяются на три типа: А, В и С. α2А-подтип обнаружен в голубом пятне, а также в коре мозга, перегородке гипоталамуса, гиппокампе и миндалине. Предполагают, что α2А-адренорецепторы принимают участие в процессах миграции, дифференцировки и созревания нейронов. α2А-адренорецепторы, локализованные на нервных окончаниях норадренергических нейронов, являются пресинаптическими ауторецепторами и угнетают высвобождение норадреналина при высокой частоте разрядной активности нейронов. α2А-адренорецепторы участвуют в регуляции кардиоваскулярной и дыхательной систем, психоэмоционального и полового поведения, вовлекаются в контроль двигательной активности.
В голубом пятне обнаружены также α1-адренорецепторы. Установлено, что данный вид рецепторов угнетает нервный эффект α2-адреноре-
цепторов. Угнетающий эффект может наблюдаться и при стимуляции
β-адренорецепторов. Активация α1-адренорецепторов ведет к высвобождению норадреналина в синаптических окончаниях аксона. Стимуляция
α2-адренорецепторов, напротив, приводит к снижению активности нервной клетки. Особенностью голубого пятна является наличие афферентных и эфферентных связей с различными отделами центральной нервной системы. Выделены три восходящие системы адренергических волокон. Первая система состоит из афферентных и эфферентных волокон, проходит вдоль пучка Фореля и достигает структур варолиева моста и среднего мозга. Большая его часть составляет центральный путь покрышки. Вторая система идет к центральному серому веществу и далее в область III и IV желудочков и сильвиева водопровода. Третья система представлена медиальным пучком переднего мозга. Наиболее мощный пучок аксонов голубого пятна направляется дорсовентрально, образуя дорсальный пучок покрышки. Часть волокон дорсального пучка покрышки присоединяется к пучку Мейнерта и достигает зрительных бугров, иннервируя его медиальные и латеральные ядра, а также ядра уздечки. Волокна голубого пятна достигают ядер гипоталамуса, в котором дают большое число коллатералей. Значительная часть их распределяется во всех областях новой коры. На уровне перегородки отделяется часть аксонов голубого пятна, которые направляются к промежуточному мозгу и каудальному отделу конечного мозга. Выявлена плотная норадренергическая иннервация базолатеральной и вентролатеральной частей миндалины, найдены терминали аксонов норадренергических нейронов голубого пятна в структурах среднего мозга, дорсальных ядрах шва и в хвостатом ядре. Голубое пятно посылает значительную часть аксонов и к мозжечку, в котором они обнаруживаются во всех его отделах. Значительная часть аксонов оканчивается возле клеток Пуркинье и в клетках молекулярного слоя. Волокна голубого пятна проецируются также в спинной мозг. Терминали аксонов клеток голубого пятна обнаружены в вентральном отделе заднего рога и в меньшей степени в дорсальной части заднего рога. Терминали голубого пятна имеются в вентральном роге, в котором аксоны голубого пятна располагаются вокруг α-мотонейронов. Высказывается мнение, что норадреналин является тормозным медиатором для данного вида мотонейронов. Широко представлены афферентные связи голубого пятна. Выявлены проекции к голубому пятну от ядра тройничного нерва, черной субстанции, дорсального и каудального отделов гипоталамуса, структур ствола мозга, дорсального ядра блуждающего нерва, ядра солитарного тракта.
Предполагается, что голубое пятно оказывает свое влияние через ретикулярную систему ствола и нейросекреторным путем. Нейросекреторный характер передачи влияний голубого пятна на центральную нервную систему можно объяснить тем, что голубое пятно обильно кровоснабжается. При этом выявлены непосредственные контакты норадренергических нейронов голубого пятна с капиллярами. Голубое пятно, имея широкие связи с различными структурами центральной нервной системы, принимает участие в регуляции многих физиологических функций. Опыты с полным или частичным выключением голубого пятна выявили его участие в регуляции суточного ритма и поведения. Выключение голубого пятна вело к уменьшению времени бодрствования и подавления фазы парадоксального сна. У животных после выключения голубого пятна в первые дни отмечались выраженный гипертонус мышц и тремор. Специфической особенностью явилась атония жевательных мышц. Развивалась гиперсаливация и задержка мочеиспускания. Через 12-15 дней после разрушения голубого пятна моторные нарушения почти полностью исчезали, но сохранялась атаксия, полифагия и полидепсия. Многочисленные данные свидетельствуют о вовлечении голубого пятна в контроль поведения. Выдвинута гипотеза о контролирующей роли голубого пятна в метаболических процессах коры головного мозга через трофическое влияние на ее структуры. Изменения функций и состояний организма при разрушении или выключении голубого пятна предположительно связывают с изменением содержания норадреналина в различных отделах мозга. Данные изменения лежат в основе нарушений механизмов памяти, обучения, поведенческих реакций и стресса. Установлено участие голубого пятна в регуляции функционального состояния слухового анализатора. Наблюдалось модулирующее влияние голубого пятна на импульсную активность нейронов пейсмекеров переднего супрахиазматического ядра гипоталамуса и формирование свободных и синхронизированных свето-темновых циклов физиологических ритмов. Это позволяет отнести голубое пятно к системе циркадного ритма. Многочисленные исследования указывают на участие голубого пятна в регуляции психоэмоционального поведения, в развитии психических расстройств и т.д. Голубое пятно принимает участие в регуляции функций сердечно-сосудистой системы, в регуляции дыхания и др.
Нейроны, содержащие дофамин, находятся в среднем мозге,образуя одно из наиболее крупных ядер вентральной части среднего мозга – черную субстанцию. Черная субстанция есть у всех позвоночных животных и человека и представлена билатеральным скоплением клеток, расположенных между основанием ножек мозга и их покрышкой. Черная субстанция состоит из сетчатой (большая) и компактной (меньшая) частей. В составе черной субстанции выделено три типа клеток. В компактной части находятся крупные клетки, встречаются также клетки средних размеров веретенообразной, треугольной или полигональной формы. В состав ретикулярной части черной субстанции входят в основном овальные и круглые нейроны малых размеров. Среди них встречаются как длинноаксонные, так и интернейроны. Исследования показали, что нейроны черной субстанции имеют полихимическую природу, но основными являются дофаминергические и ГАМКергические нейроны. По данным морфологических и электрофизиологических исследований, спонтанная и вызванная электрическая активность нейронов ретикулярной части черной субстанции определяется интеграцией возбуждающих афферентаций от субталамического ядра, ядер шва, голубого пятна, фронтальных отделов коры головного мозга, а также возбуждающих и тормозящих афферентных импульсов от неостриатума. Уровень афферентации и возбудимости нигроретикулярных нейронов обусловлен активированием или блокадой дофаминовых рецепторов. Среди них выделяют рецепторы Д1-типа, при возбуждении которых происходит активация аденилатциклазы, и рецепторы Д2-типа, при возбуждении которых аденилатциклаза либо не изменяет своей активности, либо пассивируется. Рецепторы Д1-типа обнаружены на постсинаптической мембране ГАМКергических нейронов стриатума, аксоны которых спускаются в ретикулярную часть черной субстанции. Рецепторы Д2-типа локализуются на терминалях аксонов дофаминергических нейронов черной субстанции.
Среди эфферентных связей черной субстанции выделяют нигротектальную систему, которая берет начало от нейронов ретикулярной части преимущественно ипсилатеральной черной субстанции и проецируется на таламус и передние бугры четверохолмия. Дорсальные нейроны ретикулярной части проецируются на ростральные отделы передних бугров, а ветральные – на каудальные отделы. Нигротектальный тракт, вероятно, имеет ГАМКергическую природу. Кроме того, из ретикулярной части черной субстанции берут начало нигроретикулярные волокна. Они достигают медиальной области ретикулярной формации и идут к гигантоклеточному ядру. Эта структура тесно взаимосвязана со стриарным комплексом. Нигростриарные дофаминергические волокна начинаются из компактной части черной субстанции. Повреждение черной субстанции приводит к дегенерации дофаминергических волокон, что сопровождается нарушениями тонких движений пальцев рук, развитием мышечной ригидности, тремору (болезнь Паркинсона). Функции черной субстанции традиционно связывают с регуляцией двигательных функций и поддержанием тонуса мышц. Совместно с бледным шаром черная субстанция выполняет статокинетическую функцию. При поражении черной субстанции у человека наблюдается резко выраженная бедность движений, скованность, маскообразность лица. Наблюдалось отсутствие содружественных движений, сопровождающих основной двигательный акт. Благодаря широким связям с различными ядрами ретикулярной формации ствола мозга черная субстанция оказывает влияние на вегетативные функции, принимает участие в актах глотания. Электростимуляция черной субстанции у кошек вызывала увеличение артериального давления, частоты сердечных сокращений и частоты дыхания.
В стволе мозга располагается крыша среднего мозга. Она образована передними (верхними) и задними (нижними) буграми четверохолмия. Верхние бугрыявляются сложными слоистыми и ядерными образованиями. Они участвуют в формировании зрительных и общесоматических нейронных систем. Нижние бугры – это часть слухового пути. Текто-ретикулярные волокна возникают от всех частей верхних бугров и частично от нижних. Они образуют ипсилатеральные тракты, оканчивающиеся в ретикулярной формации среднего мозга, и контрлатеральные, достигающие медиальной области ретикулярной формации продолговатого мозга и моста. В ретикулярной формации продолговатого мозга располагаются дыхательный центр, сосудодвигательный центр, центр глотания, жевания и др.
Дыхательный центр. Впервые экспериментально достаточно точное местоположение дыхательного центра в продолговатом мозге было установлено в 1885 году Н.А. Миславским. Согласно Н.А. Миславскому, дыхательный центр расположен в ретикулярной формации дна четвертого желудочка по обеим сторонам срединного шва между верхушкой и основанием писчего пера, частично кнаружи, частично кнутри от ядер подъязычного нерва. В настоящее время идет интенсивное и всестороннее изучение этой структуры, определяющей жизненно важную функцию – дыхание. По современным представлениям, дыхательный центр рассматривается в первую очередь как совокупность нейронов, активность которых синхронна с фазами дыхательного цикла. Исходя из этого признака, предложены классификации нейронов. Наиболее популярной является классификация дыхательных нейронов по Bianchi et al. (1995). Согласно этой классификации, дыхательные нейроны в соответствии с паттерном их активности подразделяются на шесть основных групп: ранние инспираторные, инспираторные с нарастающим паттерном активности, поздние инспираторные, постинспираторные, экспираторные с нарастающим паттерном активности, преинспираторные.
Дыхательные нейроны сосредоточены главным образом в пяти различных функционально-специфических отделах дыхательного центра: дорсальной дыхательной группе, вентральной дыхательной группе, пре-Бетцингеровом комплексе, комплексе Бетцингера и пневмотаксическом центре (рис. 2.3, 2.4).
Рис. 2.3. Продолговатый мозг
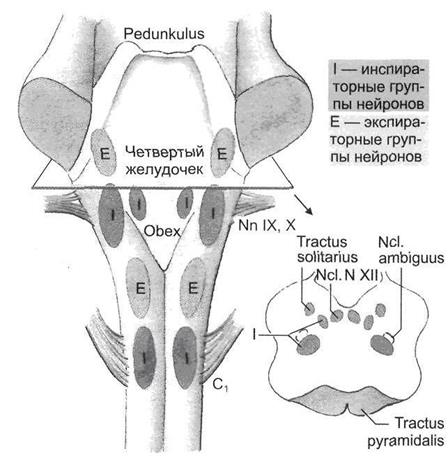
Рис. 2.4. Дыхательный центр продолговатого мозга
Дорсальная дыхательная группа нейронов. Данная группа нейронов расположена в области ядра солитарного тракта. Большинство клеток составляют инспираторные нейроны различных типов: Rα , Rβ , Р-клетки и др.
Rα-нейроны характеризуются нарастающей по ходу вдоха дыхательной активностью, а также резкой ее остановкой в момент достижения объемного порога выключения вдоха. Эти типы нейронов являются бульбо-
спинальными, их аксоны образуют перекрест на уровне каудальной части продолговатого мозга, а затем направляются к спинальным мотонейронам инспираторных мышц. Паттерн активности Rβ-нейронов напоминает таковой у Rα-нейронов, но с некоторым отличием. Активность Rβ-нейронов не обрывается в конце вдоха, а сохраняется и в начале выдоха, постепенно убывая. Установлено, что в дополнение к общему источнику драйва, получаемому от генератора центральной инспираторной активности, к Rβ-ней-
ронам приходит афферентация от рецепторов растяжения легких. Таким образом, Rβ-нейроны участвуют в реализации рефлекса Геринга-Брейера. Сравнительно недавно были описаны Р-клетки. Они получают информацию от рецепторов растяжения легких.
Большинство нейронов дорсального ядра являются бульбоспинальными (у кошек от 50 до 80%). В отношении проприобульбарных нейронов дорсальной дыхательной группы известно, что многие из них проецируются в направлении вентролатеральных отделов дыхательного центра, а также к контрлатеральному ядру солитарного тракта. Предполагают, что эти проекции играют важную роль в передаче дыхательному центру интегрирующих афферентных импульсов, оказывающих влияние на дыхание. Ядро солитарного тракта отличается многообразием нейротрансмиттеров. Его функции обеспечиваются многочисленными нейромедиаторами и нейропептидами.
Вентральная дыхательная группа. Нейроны вентральной дыхательной группы сосредоточены в двух симметричных колонках, вытянутых в рострально-каудальном направлении и расположенных в области n. ambigualis и n. retroambigualis. Вентральная дыхательная группа подразделяется на каудальный и ростральный отделы, различающиеся по функциональным характеристикам.
Каудальный отдел вентральной дыхательной группы расположен в областиn. retroambigualis. Этот отдел дыхательного центра состоит почти исключительно из бульбоспинальных экспираторных нейронов. У нейронов этой группы не обнаружено коллатералей на уровне продолговатого мозга. Предполагают, что они не имеют проекций к другим отделам бульбарного дыхательного центра. Данная группа экспираторных нейронов получает возбуждающий вход от более ростральных отделов дыхательного центра. Вероятно, активирующая афферентация поступает от экспираторных нейронов комплекса Бетцингера. Паттерн активности большинства экспираторных нейронов каудального отдела вентральной дыхательной группы характеризуется медленным нарастанием частоты импульсации на выдохе с максимумом в конце экспираторной фазы. Кроме экспираторных нейронов, в состав каудального отдела вентральной дыхательной группы входит небольшое количество инспираторных нейронов.
Ростральный отдел вентральной дыхательной группы расположен в областиn. ambigualis и прилегающих параамбигуальных структурах. Этот отдел состоит в основном из инспираторных нейронов. Здесь расположены бульбоспинальные премотонейроны инспираторных мышц, а также мотонейроны мышц гортани и некоторых мышц пищевода. В данном отделе также обнаружены проприобульбарные нейроны, которым приписывается координирующая роль в регуляции тонуса верхних дыхательных путей и инспираторных мышц. Паттерн активности инспираторных бульбоспинальных нейронов рострального отдела вентральной дыхательной группы характеризуется постепенным нарастанием частоты импульсации по ходу инспираторной фазы, аналогично паттерну активности диафрагмы и наружных межреберных мышц. В этом отделе дыхательного центра обнаружены нейроны с ранним инспираторным паттерном активности. Залпы таких нейронов характеризуются ранним началом, слегка опережающим активность диафрагмального нерва, быстрым достижением максимума с последующим снижением ее к концу инспираторной фазы. Считают, что ранние инспираторные нейроны являются источником ингибирования активности экспираторных премоторных клеток каудального отдела вентральной дыхательной группы и поздних инспираторных нейронов дорсальной дыхательной группы.
В последнее время большое значение в генерации ритма дыхания придается пре-Бетцингерову комплексу. У кошек он расположен в ростральной части n. ambigualis и вентролатеральной области ретикулярной формации каудальнее n. retrofacialis и ростральнее n. lateralis retikularis. При вне- и внутриклеточной регистрации активности нейронов здесь обнаружена область концентрации разнообразных типов дыхательных нейронов, в том числе двух типов преинспираторных нейронов, ранних инспираторных и постинспираторных нейронов. В данной структуре расположены те типы нейронов, которые согласно трехфазной теории ритмогенеза считаются необходимыми для генерации дыхательного ритма. Мнение о ведущей роли комплекса пре-Бетцингера в ритмогенезе опирается на некоторые экспериментальные данные. Так, в опытах in vitro удаление этого образования приводило к прекращению генерации дыхательного ритма в срезах ствола мозга. Локальные нарушения синаптической передачи в комплексе пре-Бетцингера также приводили к нарушению дыхательного ритма in vitro. Изучение нейронного состава комплекса пре-Бетцингера показало, что в данной структуре содержатся те типы нейронов, которые необходимы для генерации ритма в условиях in vitro: преинспираторные и пейсмекерные, а также нейроны, имеющие значения для переключения дыхательных фаз в этих условиях: пре- и постинспираторные. Следует, однако, заметить, что вывод об исключительной роли данного образования в процессе генерации ритма дыхания является преждевременным и требует экспериментального исследования в условиях in vivo.
Нейроны пре-Бетцингерова комплекса отличаются разнообразием медиаторов и их рецепторов.
Комплекс Бетцингера. Данный отдел дыхательного центра расположен в области n. retrofacialis. Большинство клеток комплекса Бетцингера относится к экспираторным нейронам. Функционально нейроны этого отдела неоднородны. Здесь обнаружены экспираторные нейроны, дающие проекции в каудальные отделы дыхательного центра и характеризующиеся нарастающим паттерном активности. Аксоны нейронов отдают множество коллатералей, большинство которых идет в направлении других вентральных отделов дыхательного центра. Экспираторные нейроны этого комплекса образуют моносинаптические тормозные проекции в направлении бульбоспинальных инспираторных нейронов дорсальной и вентральной дыхательных групп, каудальной группы ядер шва и других образований ствола. В вентральной части комплекса Бетцингера описана группа мелких нейронов с экспираторной активностью. Дендриты этих нейронов образуют широкие разветвления вблизи вентральной поверхности продолговатого мозга. Предполагают, что эта группа нейронов принимает участие в механизме центральной хеморецепции.
Вышеописанные структуры, собственно, и образуют дыхательный центр продолговатого мозга. По современным представлениям, их вполне достаточно для генерации дыхательного ритма.
На уровне организма как системы в регуляции дыхания принимают участие и другие структуры центральной нервной системы. Среди них особое место с давних пор занимают структуры варолиева моста. В настоящее время известно, что в процессе регуляции дыхания принимают участие нейроны группы А5 и А6, содержащие катехоламины, межтройничная область, а также каудальная часть ретикулярной формации моста. Наибольшее количество нейронов, разряжающихся синхронно с дыхательными фазами, обнаружено в области между парабрахиальными ядрами и ядрами Келликера-Фузе. Эта область получила название пневмотаксического центра, а совокупность находящихся здесь нейронов – понтинной дыхательной группы. Отличительной особенностью этой группы является присутствие высокого процента нейронов с пиком активности, приходящимся на момент перехода фаз дыхательного цикла. Поэтому допускается, что нейроны данной группы принимают непосредственное участие в механизмах переключения дыхательных фаз. Нейроны понтинной дыхательной группы дают широкие проекции к клеткам всех отделов бульбарного дыхательного центра (бульбоспинальным, глоточным, подъязычным мотонейронам, к спинальным мотонейронам дыхательных мышц). Получены данные, что нейроны промежуточной части ядра Келликера-Фузе контролируют постинспираторную моторную активность. Нейроны понтинной дыхательной группы получают афферентацию от различных популяций нейронов бульбарного дыхательного центра. Предполагается, что одной из важных функций нейронов этой группы является адаптация респираторных рефлексов (рефлекс Геринга-Брейера) и дыхательных рефлексов с каротидных хеморецепторов. Из этого следует, что структуры варолиева моста могут играть более значительную роль в формировании нормального и патологического паттернов дыхания, чем предполагалось
Дата добавления: 2016-07-27; просмотров: 5436;
Поиск по сайту
Узнать еще
- I. Синусы твердой оболочки головного мозга.
- II. Анатомия продолговатого мозга, ретикулярная формация
- IV. проводящие пути головного и спинного мозга
- V Патопсихологическое – при нарушении целостности мозга происходит нарушение психической деятельности
- А Нейропсихологические синдромы поражения задних отделов коры больших полушарий головного мозга.
- Абиотическая трансформация
- Абсцессы головного мозга. Хирургическое лечение
- Алгоритм зависимости длины горизонтального ствола от расхода закачиваемого газа в ПХГ.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
