ВОЗБУДИМЫЕ И НЕВОЗБУДИМЫЕ МЕМБРАНЫ
Все клеточные БМ можно разделить на возбудимые (электрогенные) и невозбудимые (неэлектрогенные). Для неэлектрогенных БМ присущ только ПП, а для возбудимых БМ присущи как ПП, так и ПД. Указанное различие обусловлено присутствием в БМ потенциалзависимых ионных каналов. В невозбудимых биологических мембранах находятся только потенциалнезависимые ионные каналы, а в возбудимых присутствуют и те, и другие. В физиологии свойства возбудимости не ограничивают мембранами, а распространяют на ткани, подразделяя их на возбудимые и невозбудимые. Считают возбудимыми такие ткани, клеточная мембрана которых обладает потенциалзависимыми ионными каналами. К ним относят нервную и мышечную ткани. Только в них под действием раздражителей возникают ПД, причем они сопровождаются целым комплексом других процессов, также развивающихся в ответ на стимуляцию (раздражение). Среди них можно отметить: изменение обмена веществ, изменение температуры, изменение электрического импеданса, а также специфические реакции, например, проведение нервных импульсов и сокращение мышечных волокон.
Способность возбудимой ткани отвечать таким образом на действия раздражителей называют возбудимостью -  . Ее количественной мерой служит интенсивность порогового раздражителя
. Ее количественной мерой служит интенсивность порогового раздражителя  , то есть самого слабого стимула, в ответ на который возникает ПД, а вслед за ним и специфическая реакция. Чем ниже интенсивность , тем выше .
, то есть самого слабого стимула, в ответ на который возникает ПД, а вслед за ним и специфическая реакция. Чем ниже интенсивность , тем выше .
Следовательно, между ними существует обратно пропорциональная зависимость:

Реакцию возбудимой ткани на раздражение называют возбуждением. У возбуждения много различных проявлений, но одним из важнейших является ПД, поскольку он проявляется при любых специфических реакциях. Плазматические мембраны нейрона или мышечного волокна возбудимы не на всем протяжении. На поверхности этих микроструктур, участки возбудимых и невозбудимых БМ чередуются, так как не везде в плазмолемму встроен потенциал зависимый ионный канал.
Существенные различия между возбудимыми и невозбудимыми БМ проявляются и из ВАХ. Для невозбудимых БМ ВАХ имеет вид:

Из ВАХ видно, что при изменении мембранного потенциала от уровня ПП (-85 мВ) до 0 ее электропроводность остается неизменной. С понижением абсолютного значения мембранного потенциала, плотность тока сквозь невозбудимые БМ пропорционально уменьшается.
ВАХ возбудимых БМ нелинейна и имеет следующий вид:
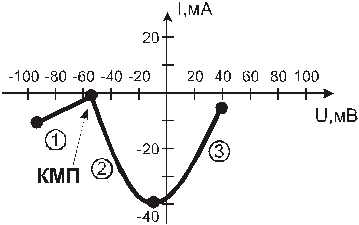
При понижении трансмембранной разности потенциалов до определенного значения, плотность тока падает до 0, а затем дальнейшее уменьшение абсолютного значения мембранного потенциала приводит к резкому нарастанию плотности тока, который к тому же изменяет свое направление (с выходящего из волокна на входящее в него). Уровень мембранного потенциала, который соответствует излому на ВАХ, называют критическим мембранным потенциалом (КМП). На рисунке КМП равен -55 мВ, что характерно для гигантского аксона кальмара. ВАХ свидетельствует о том, что деполяризация возбудимой мембраны, начиная с уровня КМП, вызывает изменение мембраной электропроводности.
ВАХ возбудимой БМ имеет N-образную форму, при этом, особое внимание заслуживает второй участок ВАХ. Его называют участком отрицательного диффузионного сопротивления (ОДС). Поскольку, при падении абсолютного значения U, I может нарастать только в том случае, если изменение напряжения сопровождается понижением сопротивления. Такая аномалия ВАХ возбудимой мембраны обусловлена присутствием в ней потенциал зависимых ионных каналов. Дело в том, что плотность трансмембранного ионного тока может определяться, в принципе, только двумя факторами:
1. Числом открытых каналов.
2. Электропроводностью каждого из них.
Известно, что электропроводность отдельного канала равна либо 0, при закрытых воротах, либо максимальному значению, порядка 1-10пОм-1, что соответствует переносу 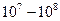 ионов за 1 секунду при открытых воротах. Не бывает промежуточных значений электропроводности отдельного канала, поэтому изменение сопротивления возбудимой мембраны зависит от количества открытых каналов при данном уровне мембранного потенциала. Сенсоры напряжения различных потенциалзависимых ионных каналов, даже если они принадлежат одному типу, например, иону
ионов за 1 секунду при открытых воротах. Не бывает промежуточных значений электропроводности отдельного канала, поэтому изменение сопротивления возбудимой мембраны зависит от количества открытых каналов при данном уровне мембранного потенциала. Сенсоры напряжения различных потенциалзависимых ионных каналов, даже если они принадлежат одному типу, например, иону  , обладают неодинаковой чувствительностью к сдвигам трансмембранной разности потенциалов, то есть, у них неодинаков порог срабатывания, поэтому в разных каналах воротные процессы включаются при различном уровне мембранного потенциала. Очевидно, что КМП соответствует такой деполяризации, при которой начинают открываться наиболее чувствительные потенциалзависимые натриевые каналы данной БМ. Начавшееся по ним движение ионов усиливает деполяризацию, которая, по мере нарастания, открывает все новые и новые потенциалзависимые натриевые каналы до тех пор, пока все они не перейдут в открытое состояние.
, обладают неодинаковой чувствительностью к сдвигам трансмембранной разности потенциалов, то есть, у них неодинаков порог срабатывания, поэтому в разных каналах воротные процессы включаются при различном уровне мембранного потенциала. Очевидно, что КМП соответствует такой деполяризации, при которой начинают открываться наиболее чувствительные потенциалзависимые натриевые каналы данной БМ. Начавшееся по ним движение ионов усиливает деполяризацию, которая, по мере нарастания, открывает все новые и новые потенциалзависимые натриевые каналы до тех пор, пока все они не перейдут в открытое состояние.
Следовательно, при изменении напряжения на аксолемме от -55 до -10 мВ (в нашем случае), потенциал зависимые натриевые каналы работают, как системы с положительной обратной связью по току. Стоит запустить в действие наиболее чувствительные компоненты такой системы, чтобы в дальнейшем вся она вовлеклась в электрогенез. Это свойство любой возбудимой мембраны. Однако, конкретные значения мембранных потенциалов, соответствующие началу и концу действия, "+" обратной связи в разных мембранах различны.
После того, как все каналы откроются, чему соответствует максимальное значение тока ионов , через определенное время мембранные ворота натриевых каналов начинают закрываться, и ток падает до нулевого значения.
РЕФРАКТЕРНОСТЬ
Процесс возбуждения сопровождается изменением возбудимости БМ. Рефрактерность - это слово, в переводе означающее "невпечатлительность". Рефрактерность - это изменение возбудимости при возбуждении. Динамику возбудимости при возбуждении во времени, можно представить в следующем виде:
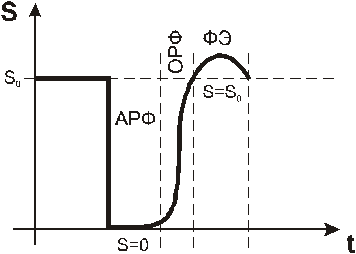
АРФ – абсолютная рефракторная фаза;
ОРФ - относительная рефракторная фаза;
ФЭ - фаза экзальтации.
На кривой выделено три участка, которые называют фазами.
Развитие возбуждения в начале сопровождается полной утратой возбудимости (S=0). Это состояние называют абсолютной рефрактерной фазой (АРФ). Оно соответствует времени деполяризации возбудимой мембраны, то есть, переход мембранного потенциала от уровня ПП до пикового значения ПД (до максимального значения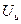 ) (см. ПД). В течение АРФ, возбудимая мембрана не может генерировать новый ПД, даже, если на нее подействовать сколь угодно сильным раздражителем. Природа АРФ состоит в том, что во время деполяризации все потенциал- зависимые ионные каналы находятся в открытом состоянии, и дополнительные стимулы (раздражители) не могут вызвать воротные процессы, так как им просто не на что действовать.
) (см. ПД). В течение АРФ, возбудимая мембрана не может генерировать новый ПД, даже, если на нее подействовать сколь угодно сильным раздражителем. Природа АРФ состоит в том, что во время деполяризации все потенциал- зависимые ионные каналы находятся в открытом состоянии, и дополнительные стимулы (раздражители) не могут вызвать воротные процессы, так как им просто не на что действовать.
АРФ сменяется относительной рефрактерной фазой (ОРФ), в течение которой возбудимость от 0 возвращается к исходному уровню (S=So). ОРФ совпадает по времени с реполяризацией возбудимой мембраны. В течение этого времени, все большее число потенциалзависимых каналов завершает воротные процессы, с которыми было связано предыдущее возбуждение. При этом каналы вновь обретают способность к следующему переходу из закрытого состояния в открытое, под действием очередного стимула. Во время ОРФ пороги возбуждения постепенно снижаются и, следовательно, возбудимость восстанавливается до исходного уровня (до Sо).
За ОРФ следует фаза экзальтации (ФЭ), для которой характерно повышенная возбудимость (S>So). Она, очевидно, связана с изменениями свойств сенсора напряжения во время возбуждения. Предполагается, что за счет конформационных перестроек белковых молекул изменяется их дипольный момент, что приводит к повышению чувствительности сенсора напряжения к сдвигам мембранной разности потенциалов, то есть, критический мембранный потенциал как бы приближается к ПП.
Разные мембраны имеют неодинаковые продолжительности каждой фазы. Так, например, в скелетных мышцах АРФ в среднем длится 2,5 мс, ОРФ - порядка 12 мс, ФЭ - 2 мс. Миокард человека отличается очень долгой АРФ, равной 250-300 мс, что обеспечивает четкую ритмичность сердечных сокращений. Различие во времени каждой фазы объясняется тем, какие каналы ответственны за этот процесс. В тех мембранах, где возбуждаемость обеспечивается натриевыми каналами, рефрактерные фазы наиболее быстротечны, и ПД имеет наименьшую продолжительность. Если же за возбудимость ответствены кальциевые каналы, то рефрактерные фазы затягиваются до секунд. В мембране миокарда человека присутствуют и те, и другие каналы ( и 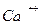 ), вследствие чего длительность рефракторных фаз занимает промежуточное значение.
), вследствие чего длительность рефракторных фаз занимает промежуточное значение.
РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ
Возбудимая мембрана относится к нелинейным и активным средам. Активной называют такую среду, которая генерирует электромагнитную энергию под действием приложенного к ней электромагнитного поля. Способность к БЭГ (к образованию ПД) отображает активный характер возбудимости мембраны. Активный характер проявляется также и в наличии участка ОДС на ее ВАХ. Это же свидетельствует о нелинейности возбудимой мембраны, поскольку, отличительным признаком нелинейности среды служит нелинейная функция, зависимость потоков от сил, которые их вызывают. В нашем случае - это зависимость ионного тока от трансмембранного напряжения. Применительно к электрическому процессу в целом, это означает нелинейную зависимость тока от напряжения.
Нервные и мышечные волокна, будучи генераторами ЭМЭ (электромагнитной энергии), обладают и пассивными электрическими свойствами. Пассивные электрические свойства характеризуют способность живых тканей поглощать энергию внешнего ЭМП (электромагнитного поля). Эта энергия затрачивается на их поляризацию, и она характеризуется потерями в тканях. Потери в живых тканях приводят к затуханию ЭМП, то есть, говорят о декременте. Закономерности затухания ЭМП идентичны для потенциалов, приложенных извне, и генерируемых самими живыми тканями (ПД). Степень декремента (затухания) зависит от сопротивления и емкости ткани. В электронике, сопротивление и емкость (индуктивность) называют пассивными свойствами электрических цепей.
Допустим, что в какой-то точке БМ потенциал мгновенно возрос до величины  . В результате затухания во времени потенциал будет уменьшаться по exp закону:
. В результате затухания во времени потенциал будет уменьшаться по exp закону:

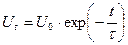
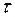 - постоянная времени затухания, то есть, время, в течение которого амплитуда уменьшается в e раз (37 %).
- постоянная времени затухания, то есть, время, в течение которого амплитуда уменьшается в e раз (37 %).
Постоянная времени зависит от пассивных свойств нервных или мышечных волокон:

Так, например, для гигантского аксона кальмара, Rн составляет примерно 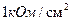 , а
, а 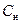 равно примерно
равно примерно  , следовательно, равна примерно 1 мс.
, следовательно, равна примерно 1 мс.
Угасание потенциала происходит не только с течением времени в точке его возникновения, но и также, при распределения потенциала вдоль БМ, по мере удаления от этой точки. Такой декремент является функцией не времени, а расстояния:

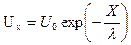
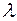 - постоянная длины, то есть это расстояние, на котором
- постоянная длины, то есть это расстояние, на котором 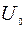 уменьшается в
уменьшается в  раз.
раз.
Декремент потенциала вдоль БМ происходит достаточно быстро в обе стороны от того места, где возник скачок мембранного потенциала. Распределение электрического потенциала на БМ устанавливается практически мгновенно, так как скорость распределения ЭМП близка к скорости распространения света ( 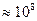 м/с). С течением времени, потенциал падает во всех точках волокна (мышечного или нервного). Для длительно существующих сдвигов мембранного потенциала, постоянная длины вычисляется по формуле:
м/с). С течением времени, потенциал падает во всех точках волокна (мышечного или нервного). Для длительно существующих сдвигов мембранного потенциала, постоянная длины вычисляется по формуле:

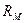 - погонное сопротивление мембраны (
- погонное сопротивление мембраны ( 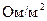 );
);
 - сопротивление цитоплазмы (Ом);
- сопротивление цитоплазмы (Ом);
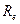 - сопротивление межклеточной среды (Ом).
- сопротивление межклеточной среды (Ом).
При коротких импульсах, как ПД, необходимо учитывать емкостные свойства БМ. Из экспериментов установлено, что емкость БМ вносит искажение в эту формулу. С учетом поправки, постоянная длины для ПД, оценивается величиной .

Чем больше , тем слабее декремент потенциала вдоль мембраны. Так, в гигантском аксоне кальмара примерно равна 2,5 мм. У больших волокон составляет примерно 10-40 их диаметров.
Таким образом, и являются основными параметрами, которые характеризуют кабельные свойства БМ. Они количественно определяют декремент потенциала, как во времени, так и в пространстве. Для уяснения механизмов распределения возбуждения, особо важное значение имеет волокон. Анализ кабельных свойств нервных и мышечных, свидетельствует о их крайне низкой электропроводности. Так называемый аксон, диаметром 1 микрон и длиной 1 м, имеет сопротивление 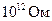 . Поэтому, в невозбудимой мембране всякий сдвиг мембранного потенциала быстро затухает в окрестности того места, где он возник, что полностью соответствует кабельным свойствам.
. Поэтому, в невозбудимой мембране всякий сдвиг мембранного потенциала быстро затухает в окрестности того места, где он возник, что полностью соответствует кабельным свойствам.
Возбудимым мембранам также присущ декремент потенциала, по мере удаления от места возникновения возбуждения. Однако, если затухающий потенциал достаточен для включения воротного процесса потенциал зависимых ионных каналов, то на удалении от первичного очага возбуждения возникает новый ПД. Для этого должно соблюдаться условие:

Регенерированный ПД также будет распределяться с декрементом, но, угасая сам, он возбудит последующий участок волокна, и этот процесс повторяется многократно:
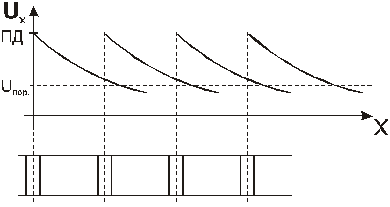
В силу огромной скорости декрементного распространения потенциала электроизмерительные приборы не способны зарегистрировать угасание каждого предыдущего ПД на последующих участках БМ. Вдоль всей возбудимой мембраны, при распределении по ней возбуждения, приборы регистрируют только одинаковые по амплитуде ПД. Распределение возбуждения напоминает горение бикфордова шнура. Создается впечатление, будто электрический потенциал распределяется по БМ без декремента. На самом деле, бездекрементное движение ПД по возбудимой мембране, является результатом взаимодействия двух процессов:
1. Декрементного распространения потенциала от предыдущего ПД.
2. Генерация нового ПД. Этот процесс называют регенерацией.
Первый из них протекает на несколько порядков быстрее, чем второй, поэтому, скорость проведения возбуждения по волокну тем выше, чем реже приходится ретранслировать (регенерировать) ПД, что, в свою очередь, зависит от декремента потенциала вдоль БМ ( ). Волокно, обладающее большей , быстрее проводит нервные импульсы (импульсы возбуждения).
В физиологии принят и иной подход для описания распределения возбуждения по нервным и мышечным волокнам, которое не противоречит рассмотренному выше. Этот подход был разработан Германном, и его называют методом локальных токов

1 - возбудимый участок;
2 - невозбудимый участок.
Согласно этой теории, между возбужденным и невозбужденным участками волокна течет электрический ток, так как внутренняя поверхность первого из них обладает положительным потенциалом относительно второго, и между ними существует разность потенциалов. Токи, возникающие в живых тканях вследствие возбуждения, называются локальными, так как распределяются на незначительное расстояние от возбужденного участка. Их ослабление обусловлено затратами энергии на заряд мембраны и на преодоление сопротивления цитоплазмы волокна. Локальный ток служит раздражителем для покоящихся участков, которые непосредственно прилегают к месту деполяризации (возбуждения). В них развивается возбуждение, а значит и новая деполяризация. Она приводит к установлению разности потенциалов между вновь деполяризированными и покоящимися (последующими) участками волокна, вследствие чего, возникает локальный ток в следующем микроконтуре, следовательно, распределение возбуждения представляет собой многократно повторяющийся процесс.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА СКОРОСТЬ
РАСПРЕДЕЛЕНИЯ ВОЗБУЖДЕНИЯ
Скорость распределения возбуждения возрастает по мере понижения сопротивления цитоплазмы и емкости клеточной мембраны, так как 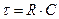 . Сопротивление определяется по формуле:
. Сопротивление определяется по формуле:

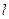 - длина нервного волокна;
- длина нервного волокна;
S - сечение нервного волокна;
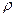 - удельное сопротивление цитоплазмы.
- удельное сопротивление цитоплазмы.
Толстые волокна обладают низким сопротивлением, и, вследствие этого, быстрее проводят возбуждение. Так, в ходе эволюции, некоторые животные приобрели способность к быстрой передаче нервных импульсов, за счет образование в них толстых аксонов, путем слияния многих мелких в одно крупное. Примером служит гигантское нервное волокно кальмара. Его диаметр достигает 1-2 мм, тогда, как обычное нервное волокно имеет диаметр от 1-10 микрон.
Эволюция животного мира привела и к использованию другого пути повышения скорости передачи нервной импульсации, то есть, уменьшению емкости плазматической мембраны аксона (аксолемма). В результате появились нервные волокна, покрытые миелиновой оболочкой. Они называются мякотными или миелиновыми. Миелиновая оболочка образуется в процессе "наматывания" на аксон клеток. Оболочка представляет собой много мембранную систему, включающую от нескольких десятков, до 200 элементов клеточных мембран, которые прилегают друг к другу и, при этом, внутренний их слой образует плотный электрический контакт с аксолеммой. Толщина всей миелиновой оболочки сравнительно невелика (1 микрон), но это достаточно для значительного понижения емкости мембраны. Так как миелин является хорошим диэлектриком (удельное сопротивление миелиновой оболочки составляет примерно 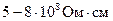 ), емкость мембраны миелинового аксона примерно в 200 раз меньше емкости аксона без мякотного волокна, то есть, примерно 0,005 и
), емкость мембраны миелинового аксона примерно в 200 раз меньше емкости аксона без мякотного волокна, то есть, примерно 0,005 и 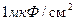 соответственно.
соответственно.
Диффузия ионов через миелиновую оболочку практически невозможна, кроме того, в участках аксона, покрытых ею, отсутствует потенциал зависимые ионные каналы. В этой связи, в мякотном нервном волокне, места генерации ПД сосредоточены только там, где миелиновая оболочка отсутствует. Эти места в мембране миелинового аксона называются перехватами Ранвье или активными узлами. От перехвата к перехвату нервные импульсы проводятся за счет декрементного распределения электромагнитного поля (движение локальных токов). Расстояние между соседними перехватами составляет в среднем 1 мм, но оно сильно зависит от диаметра аксона. Так, например, у животных эта зависимость выражается следующим образом:
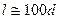 .
.
Перехваты Ранвье занимают примерно 0,02 % общей длины нервного волокна. Площадь каждого из них около 20 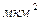 .
.
Время проведения возбуждения между соседними активными узлами составляет примерно 5-10 % длительности ПД. В этой связи, сравнительно большой путь (около 1 мм) между следующими друг за другом участками ретрансляции ПД обеспечивает высокую скорость проведения нервного импульса. Необходимо отметить, что локальные токи,
достаточные для регенерации ПД, могут даже протекать через 2-3 последовательно расположенные перехвата Ранвье. Более частое, чем необходимо для обеспечения нормального распределения возбуждения, расположение активных узлов в мякотных аксонах, служит повышением надежности нервных коммуникаций в организме. У гомойотерных животных надежность выше, чем у пойкилотерных (животных с переменной температурой). В без-мякотных аксонах ретрансляция ПД происходит значительно чаще. Там генераторы ПД расположены вдоль всей длины волокна, в непосредственной близости друг от друга (около 1 микрона). Это обусловлено сравнительно низкой скоростью проведения возбуждения по мембранам мышечных и нервных волокон, которые не покрыты миелиновой оболочкой. В отличие от них, миелиновые аксоны за счет малой емкости между участками перехватов Ранвье, приобрели высокую скорость передачи нервных импульсов (до 140 м/с).
Вследствие относительно большой протяженности участков аксона между соседними активными узлами, проведение нервного импульса в мякотном нервном волокне, происходит как бы скачками, и поэтому его называют сальтоторным. Сальтоторное проведение обеспечивает существенную экономию энергии. Так, например, потребление 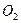 при нем в 200 раз меньше, чем при непрерывном распределении нервных импульсов по безмякотным аксонам. Наибольшая скорость распространения возбуждения наблюдается в мякотных аксонах, диаметр которых составляет примерно 10-15 микрон, а толщина миелиновой оболочки достигает 30-50 % общего диаметра волокна. Скорость проведения нервных импульсов в миелиновых аксонах пропорциональна их диаметру, тогда как в безмякотных аксонах скорость проведения возбуждения пропорциональна корню квадратному из диаметра.
при нем в 200 раз меньше, чем при непрерывном распределении нервных импульсов по безмякотным аксонам. Наибольшая скорость распространения возбуждения наблюдается в мякотных аксонах, диаметр которых составляет примерно 10-15 микрон, а толщина миелиновой оболочки достигает 30-50 % общего диаметра волокна. Скорость проведения нервных импульсов в миелиновых аксонах пропорциональна их диаметру, тогда как в безмякотных аксонах скорость проведения возбуждения пропорциональна корню квадратному из диаметра.
Дата добавления: 2020-11-18; просмотров: 1322;
Поиск по сайту
Узнать еще
- Активно-возбудимые среды. Автоволновые процессы в сердечной мышце
- БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
- Вопрос 17. Мембраны, их молекулярная структура
- ЗАКОНЫ ДЕЙСТВИЯ ПОСТОЯННОГО ТОКА НА ВОЗБУДИМЫЕ ТКАНИ.
- ЛОКАЛЬНЫЙ ПОТЕНЦИАЛ. ОЦЕНКА ПРОНИЦАЕМОСТИ КЛЕТОЧНОЙ МЕМБРАНЫ
- Мембранный потенциал покоя – это разность потенциалов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя.
- Модели механизмов массопереноса растворенных веществ через полупроницаемые мембраны при обратном осмосе

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
