ТРАНСПОРТ ВЕЩЕСТВ В ОРГАНИЗМЕ. БИОМЕМБРАНОЛОГИЯ
СТРУКТУРА И ФУНКЦИИ БИОЛОГИЧЕСКИХ МЕМБРАН
Для организма, как для открытой системы, характерен обмен с окружающей средой энергией, веществом и информацией. Необходимым условием обмена веществ является их транспорт внутри биологической системы. Все вещества, из которых состоит организм, перемещаются в нем с потоком крови и лимфы, они преодолевают значительные расстояния, обеспечивая обмен между органами и тканями. В ходе обмена между кровью и тканями, вещества проникают сквозь стенки кровеносных сосудов. Внутри клеток происходит непрерывный транспорт веществ через мембраны органелл. Заболевания, при которых нарушен транспорт веществ в организме, связаны, прежде всего, с различными нарушениями молекулярной организации биологических мембран – (БМ).
Термин "мембрана" используют в биологии, обозначая им клеточную границу, которой свойственна полупроницаемость, то есть легкость проникновения сквозь нее одних веществ при невозможности преодоления ее другими.
Каждая клетка окружена наружной мембраной, которая называется плазматической мембраной или плазмалеммой. В настоящее время клетку стали воспринимать, как обширную сеть мембранных систем, составляющих важнейший элемент клеточной организации; соотношение между плазмалеммой и внутриклеточной мембраной неодинаково в различных клетках. Так, в клетках хрусталика глаза нет никаких других мембран, кроме плазматических, тогда как в почечных клетках (гепатоцитах) площадь плазматических мембран составляет порядка 6% от поверхности, занимаемой всеми мембранами клетки. Установлено, что по мере увеличения отношения сумм площади мембран к объему клетки повышается интенсивность обменных процессов в этой клетке.
Электронная микроскопия клеточных мембран не позволяет их увидеть, так как они состоят из тех же химических элементов, что и цитоплазма. Для получения четкой электронограммы клетки ее мембраны контрастируют. Для этого производят осаждение на них вольфрама, осмия и других элементов, которые хорошо поглощают и рассеивают электроны. На таких препарированных клетках любая БМ выглядит трехслойной: между двумя темными полосами располагается светлое пространство.
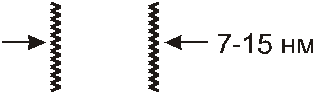
Следовательно, компоненты промежуточной части БМ слабо связывают вещества, входящие в состав красители. Суммарная толщина трехслойной структуры может изменяться от 7 до 15 нм, причем, разная величина присуща различным клеточным мембранам. Во многих БМ наблюдается асимметрия трехслойной организации, то есть темные полосы различны по ширине и по плотности.
Все клеточные мембраны построены в основном из липидов, белков (протеинов) и углеводов. При этом, углеводы образуют соединения с белками (гликопротеины) и липидами (гликолипиды). Органические вещества образуют соли с различными ионами, которые присутствуют в виде водных растворов внутри мембранных каналов.
Структурной основой БМ служит бимолекулярный слой. Его образование обусловлено особенностями взаимодействия с водой мембранных липидов, среди которых преобладают фосфолипиды. В молекуле фосфолипидов условно выделяются 3 части:
- головка;
- тело;
- пара длинных хвостов.
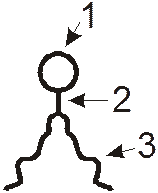
Головка (1) образована одним из таких соединений, как холин, серин, треонин, инозин, эталомин. Головка содержит и остаток фосфорной кислоты.
Тело (2) состоит из молекулы глицерина или сфингозина.
Пара длинных хвостов (3) содержит углеводные цепи жирных кислот. При этом преобладают не разветвленные цепи с четным числом атомов С (обычно от 14 до 24). При этом в каждой из цепей содержится до шести двойных связей. Чаще всего встречаются стеариновая, пальмитиновая, оленовая, линолевая, линоленовая, арахидоновая кислоты. Чем больше в мембранах ненасыщенных жирных кислот, тем меньше ее жесткость и выше проницаемость для веществ. Так, по мере увеличения числа двойных связей в неполярных цепях фосфолипидов, удлиняются промежутки между головками двух соседних молекул.
Головка фосфолипида либо электронейтральна, либо несет отрицательный заряд. Образующие ее молекула, а также глицерин или сфингозин - гидрофильны, тогда как жирно-кислотные хвосты - гидрофобны. В этой связи, в воде молекулы фосфолипидов самопроизвольно выстраиваются так, что их гидрофобные углеводородные цепи оказываются укрытыми от воды, а головки вступают во взаимодействие с водой. В результате создается конструкция, поперечник которой включает две молекулы фосфолипидов, повернутых друг к другу жирно-кислотными хвостами и обращенные к обеим наружным поверхностям гидрофильными головками. Графически это можно представить так:

Так образуется сплошной бимолекулярный фосфолипидный слой (бислой), который и служит каркасом биологической мембраны. Для создания 1 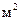 поверхности БМ достаточно 1 мг фосфолипида. При этом, на каждый квадратный микрон клеточной мембраны, приходится примерно
поверхности БМ достаточно 1 мг фосфолипида. При этом, на каждый квадратный микрон клеточной мембраны, приходится примерно 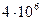 липидных молекул, то есть, по 2 млн в каждом из моно слоев бислойной структуры.
липидных молекул, то есть, по 2 млн в каждом из моно слоев бислойной структуры.
Наряду с фосфолипидами, в состав биологических мембран могут входить и другие жиры, причем характерным свойством большинства из них является наличие пары углеводородных хвостов. Одноцепочные липиды встречаются крайне редко. В их присутствии клеточные мембраны имеют тенденцию к разрушению. С этим связана причина гибели людей при укусах тех видов змей, в яде которых имеется особый фермент - фосфолипаза  . Он превращает двухцепочные фосфолипиды в одноцепочный липид - лизолецитин, в результате чего мембраны клеток разрушаются. Другой фермент - фосфолипаза
. Он превращает двухцепочные фосфолипиды в одноцепочный липид - лизолецитин, в результате чего мембраны клеток разрушаются. Другой фермент - фосфолипаза 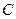 , выделяемый некоторыми микроорганизмами (например, холерным виброном), разрушает БМ, "откусывая" полярные головки фосфолипидов.
, выделяемый некоторыми микроорганизмами (например, холерным виброном), разрушает БМ, "откусывая" полярные головки фосфолипидов.
Особое место среди мембранных липидов занимает холестерин. По мере повышения содержания холестерина в БМ площадь, занимаемая фосфолипидами, сокращается до тех пор, пока на одну молекулу холестерина не будет приходиться две молекулы фосфолипида. Эффект сокращения площади, приходящийся на одну молекулу фосфолипида, обусловлен тем, что изменяется наклон его углеводородных цепей к поверхности бислоя. По мере увеличения содержания холестерина в БМ фосфолипиды стремятся встать перпендикулярно мембранной поверхности. В результате этого укладка фосфолипидов приобретает большую компактность и мембрана уплотняется. В результате, она становится более вязкой и менее проницаемой для многих веществ (например, глюкозы, ионов воды и т. д.). Содержание холестерина в клетках зависит от общего холестеринового обмена в организме, который, в свою очередь, сильно подвержен влиянию пищевого рациона.
МЕМБРАННЫЕ БЕЛКИ
В липидный каркас клеточных мембран встроены белковые компоненты (протеины). На каждую клетку в среднем приходится около 10 пг мембранных белков (МБ). Различают периферические и собственные (интегральные) белки биологических мембран. Белки первого типа располагаются на поверхности липидного бислоя. Здесь связь между липидами и белками осуществляется за счет электростатического взаимодействия между противоположно заряженными полярными группами этих веществ. Интегральные белки взаимодействуют своими гидрофобными областями с углеводородными цепочками липидов только за счет Ван-дер-Ваальсовых сил. У интегральных белков все гидрофильные области спрятаны внутрь молекулы, а наружу направлены неполярные группы. В этой связи, собственные белки полностью или частично погружаются в биологическую мембрану, при этом крупные белковые молекулы могут пронизывать БМ насквозь.
Наиболее характерным типом вторичных структур интегральных белков является спираль. Причем, на участках, окруженных липидами, преобладает 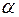 -спираль, у которой все аминокислотные остатки - гидрофобные. Реже встречается
-спираль, у которой все аминокислотные остатки - гидрофобные. Реже встречается 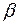 - спираль, представляющая собой полое микротрубчатое образование, на наружных стенках которого сосредоточены гидрофобные аминокислотные остатки, а внутрь полости ориентированы гидрофильные группы. Предполагается, что белки, имеющие структуру - спирали, образуют ионные каналы в клеточных мембранах.
- спираль, представляющая собой полое микротрубчатое образование, на наружных стенках которого сосредоточены гидрофобные аминокислотные остатки, а внутрь полости ориентированы гидрофильные группы. Предполагается, что белки, имеющие структуру - спирали, образуют ионные каналы в клеточных мембранах.
Функции мембранных белков весьма разнообразны. Среди них можно выделить следующие основные:
1. Обеспечение транспортных свойств БМ (образование мембранных каналов, работа в качестве переносчиков определенных веществ, участие в системах активного транспорта);
2. Белки несут функцию структурного компонента БМ, тем самым, они усиливают прочность липидного каркаса.
3. Большим разнообразием различаются мембранные протеины, катализирующие химические реакции. Причем белки-ферменты, встроенные в БМ, действуют активнее, чем при их пребывании в растворе. Установлено, что основная часть всех биохимических реакций, протекает на клеточных мембранах.
4. Важным функциональным типом МБ являются белки-рецепторы. Они столь разнообразны, сколь разнообразны стимулы, которые воспринимаются рецепторными белками. Так, например, восприятие света, обеспечивается пигментами, в состав которых, наряду с белками, входят хромофорные группы с определенным спектром поглощения. Таким пигментом является родопсин, который обеспечивает реакции на свет палочек или колбочек.
5. Белки, присутствующие в мембранах любой клетки, осуществляют ее взаимодействие с окружающей средой, а также обмен информацией между органеллами внутри клетки.
6. Белки, образующие антитела, способны связывать специфический антиген (белок бактерии, вируса, токсина и т. д.) и вызывать иммунный ответ клетки, в плазмолемме которой белки сосредоточены.
В различных мембранах белки распределены среди липидов по-разному. В плазмолемме их распределение довольно равномерное. Для специализированных внутриклеточных мембран, характерно неоднородное распределение белков в БМ.
Различия между клеточными мембранами разных типов, между участками БМ и даже между сторонами одной и той же мембраны привели к выводу о гетерогенности БМ. Под гетерогенностью понимают разнородность их структур и функциональных свойств. Особенности молекулярных структур клеточных мембран и предопределяют их физические и физико-химические свойства.
ФИЗИЧЕСКИЕ И ФИЗИКО-ХИМИЧЕСКИЕ
СВОЙСТВА БМ
В живых клетках БМ представляют собой жидкокристаллические структуры. Структуры жидкого кристалла образуют как липидные, так и белковые молекулы. Молекулярной организации клеточных мембран, как жидкого кристалла, характерно состояние с высокой степенью упорядоченности и текучести (лабильности).
БМ присущи такие свойства:
1. Значительная прочность на разрыв.
2. Упругость (эластичность).
3. Поверхностное натяжение.
4. Вязкость.
5. Электрострикция.
6. Флексоэлектрострикция.
Два последних свойства обусловлены наличием повторного заряда на БМ.
ПОВЕРХНОСТНЫЙ ЗАРЯД НА КЛЕТОЧНОЙ МЕМБРАНЕ (КМ)
Он создается полярными головками фосфолипидов, гликопродеидами (главным образом, карбоксильными группами сиаловой кислоты и аминокислотными остатками), а также, гликолипидами. За счет этих веществ поверхность БМ заряжена отрицательно. Поверхностный заряд плазмолеммы играет важную роль в межклеточных взаимодействиях. Он способствует стабильности мембранных структур, а также связыванию ионов, находящихся в межклеточной среде. От поверхностного заряда плазмолеммы зависит ионный состав предмембранных слоев, что оказывает влияние на многие внутриклеточные обменные процессы.
Существование заряженных групп на БМ приводит к образованию двойного диффузионного электронного слоя, в котором фиксированный отрицательный заряд клеточной поверхности уравновешен положительным зарядом, который создается межклеточной средой за счет положительных ионов. Подвижность ионов не одинакова в различных условиях. Разность потенциалов между неподвижными и подвижными частями двойного электронного слоя называют электрокинетическим потенциалом, или дзета- потенциалом. Толщина двойного электрического слоя и дзета-потенциала зависит от природы электролита и концентрации ионов (в межклеточной среде). Так, при уменьшении концентрации 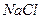 в межклеточной среде в 200 раз толщина двойного электрического слоя возрастает в 5 раз. При высокой концентрации электролита, дзета потенциал стремиться к нулю. Когда в межклеточной среде присутствуют двухвалентные катионы, избыток положительного заряда может стать настолько велик, что дзета потенциал изменит свой знак. Снижение дзета потенциала и, тем более, изменение его знака на противоположный, вызывает слипание плазматических мембран соседних клеток. Это проявляется при избытке ионов в межклеточной среде, а также, при перестройках клеточных мембран, приводящих к сдвигам дзета потенциала. Подобный эффект имеет свои особенности у клеток крови, в которых дзета потенциал форменных элементов (эритроцитов) может падать за счет нарушения не только солевого, но и белкового состава кровяной плазмы. Это явление лежит в основе изменения скорости осаждения эритроцитов (СОЭ), измерение которых широко используется в диагностических целях.
в межклеточной среде в 200 раз толщина двойного электрического слоя возрастает в 5 раз. При высокой концентрации электролита, дзета потенциал стремиться к нулю. Когда в межклеточной среде присутствуют двухвалентные катионы, избыток положительного заряда может стать настолько велик, что дзета потенциал изменит свой знак. Снижение дзета потенциала и, тем более, изменение его знака на противоположный, вызывает слипание плазматических мембран соседних клеток. Это проявляется при избытке ионов в межклеточной среде, а также, при перестройках клеточных мембран, приводящих к сдвигам дзета потенциала. Подобный эффект имеет свои особенности у клеток крови, в которых дзета потенциал форменных элементов (эритроцитов) может падать за счет нарушения не только солевого, но и белкового состава кровяной плазмы. Это явление лежит в основе изменения скорости осаждения эритроцитов (СОЭ), измерение которых широко используется в диагностических целях.
Дзета потенциал, измеряемый у различных клеток, изменяется в пределах от -10 до -30 мВ. Его величина спадает по exp закону, с увеличением расстояния от поверхности БМ. Дзета потенциал оказывает существенное влияние на размеры межклеточных пространств, и противодействует силам их притяжения. Наименьшее расстояние между клетками составляет порядка 10-20 нм. При таком расстоянии существует энергетическая "яма" во взаимодействии кулоновских и Ван-дер-Ваальсовских сил. В большинстве тканей расстояние между клетками составляет от 0,1 до 1 микрона.
Липидная часть БМ обладает свойствами диэлектрика. Относительная диэлектрическая проницаемость ( 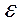 ) гидрофобной зоны составляет порядка 2-2,2, тогда, как в гидрофильной части
) гидрофобной зоны составляет порядка 2-2,2, тогда, как в гидрофильной части 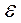 гораздо выше, и составляет около 10-20. Емкость полярных головок липидов достигает примерно 30
гораздо выше, и составляет около 10-20. Емкость полярных головок липидов достигает примерно 30 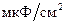 , а жирно-кислотные хвосты имеют емкость около 0,5-0,9 . Общая емкость БМ составляет примерно 1 .
, а жирно-кислотные хвосты имеют емкость около 0,5-0,9 . Общая емкость БМ составляет примерно 1 .
Благодаря существованию в БМ заряженных групп, ей присуща ярко выраженная электрострикция, то есть, по мере повышения трансмембранной разности потенциалов, БМ сжимается, что приводит к утоньшению гидрофобной зоны мембраны и, соответственно, к увеличению мембранной емкости. Возрастание дзета потенциала сопровождается увеличением клеточной поверхности.
МЕХАНИЧЕСКИЕ СВОЙСТВА БМ
Механические свойства КМ весьма своеобразны. Величина модуля Юнга в поперечном направлении достигает  . Вместе с тем, устойчивость к деформации сдвига примерно на 4 порядка меньше. Сила, достаточная для разрушения БМ на участке в 1 мкм, оценивается величиной примерно
. Вместе с тем, устойчивость к деформации сдвига примерно на 4 порядка меньше. Сила, достаточная для разрушения БМ на участке в 1 мкм, оценивается величиной примерно 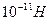 . Значительный вклад в повышение упругости и прочности мембраны вносят не только интегральные, но, также, довольно густая цепь периферических белков.
. Значительный вклад в повышение упругости и прочности мембраны вносят не только интегральные, но, также, довольно густая цепь периферических белков.
ВЯЗКОСТЬ БМ
В живой клетке БМ находятся в жидкокристаллическом состоянии. Жидкокристаллическому состоянию присущи текучесть и определенная вязкость. Величина вязкости была измерена с помощью ЭПР (эл. парамагнитного резонанса) и ДСК (диффузионно - сканирующей микрокаллометрией). Результаты измерения вязкости разными методами совпали. При этом вязкость БМ изменилась от 0,03 до 0,1 Па*с (30-100 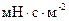 ) или 30-100 сП (сантипуаз). Это примерно в 30-100 раз больше, чем вязкость воды и примерно такая же, как вязкость оливкового или подсолнечного масла. При изменении температуры молекулярного состава и других параметров биологических мембран, изменяется ее вязкость, вследствие чего происходят фазовые изменения в структуре мембраны, то есть, осуществляется переход жидкий кристалл - твердый кристалл.
) или 30-100 сП (сантипуаз). Это примерно в 30-100 раз больше, чем вязкость воды и примерно такая же, как вязкость оливкового или подсолнечного масла. При изменении температуры молекулярного состава и других параметров биологических мембран, изменяется ее вязкость, вследствие чего происходят фазовые изменения в структуре мембраны, то есть, осуществляется переход жидкий кристалл - твердый кристалл.
Одним из важных достоинств температурного гомеостаза является поддержание БМ в жидкокристаллическом состоянии, что обеспечивает стабильность транспорта веществ через нее. Жизнеспособность организма страдает как при понижении, так и при повышении мембранной проницаемости, которая, в свою очередь, зависит от вязкости БМ.
Дата добавления: 2020-11-18; просмотров: 686;
Поиск по сайту
Узнать еще
- I. 3. Функции минеральных веществ плазмы крови
- III. Биогеохимические циклы элементов и веществ (на примере основных биогенных элементов: углерод, азот, фосфор) и их количественные характеристики.
- III. Характеристики основных классов загрязняющих веществ в водной среде.
- IV. Основные источники поступления загрязняющих веществ в водную среду.
- VIII. 2. Большой круговорот вещества и роль в нем человека
- А.3. Транспорт и монтаж сборных элементов
- Абиотическая миграция вещества
- Авіаційний транспорт

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
